ABSTRACT
FLORA OF VIETNAM 15
ASCLEPIADACEAE R. Br. 1810
(3 subfamilies, 6 tribes, 49 genera, 143 species;
Information on 1 genus and 13 species is not available)
INTRODUCTION
The family Asclepiadaceae R. Br. comprises approximately 250 genera and more than 2000 species, and is widely distributed in tropical and subtropical. In Vietnam, 49 genera and 143 species of Asclepiadaceae have been recognized. The family comprises approximately 60 useful species (35 genera) used as medicinal, ornamental and edible plants.
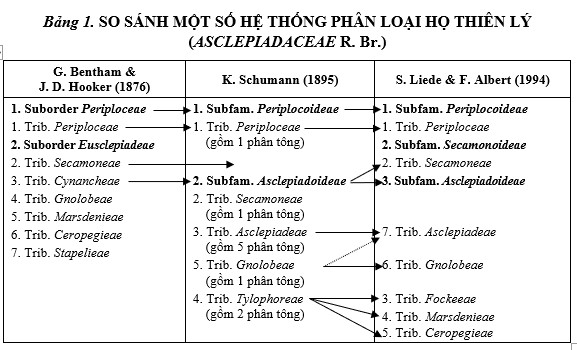
The family Asclepiadaceae was established in 1810 by R. Brown, separating it from family Apocynaceae Juss. (1789) and further divided into 3 tribes and 37 genera. Many taxonomists such as C. L. Blume (1826), J. Lindley (1836), H. G. Reichenbach (1837) supported this classification. However, G. Bentham & J. D. Hooker (1876) divided the family into 2 subfamilies, 7 tribes, and 146 genera. Another system by K. Schumann (1895) divided the family into 2 subfamilies, 4 tribes, and 10 subtribes. This study also mentioned the phylogeny of Asclepiadaceae. Further, F. R. R. Schulter (1905), A. A. Bullock (1956), and J. Hutchinson (1973) divided the family into 3 subfamilies. Lately, S. Liede & F. Albers (1994) divided Asclepiadaceae into 3 subfamilies and 7 tribes. In the present work, this system of classification has been followed to arrange taxa of Asclepiadaceae in Vietnam.
In Vietnam, J. Loureiro (1790) was the first to study Asclepiadaceae in detail. He divided the family into 17 genera and 26 species. In 1912, J. Costantin again classified the family comprising 104 species, 38 genera. Thereafter, many other significant works on Asclepiadaceae were published by the Vietnamese scientists (Le Kha Ke (1970), Pham Hoang Ho (1972, 1993, 2000), Tran The Bach (2001, 2002, 2003, 2004, 2005,…)… Publications related to medicinal plants were also came out during this period (A. Petelot (1953), Do Tat Loi (1995), Vo Van Chi (1997, 2012),…). However, a taxonomical monograph of Asclepiadaceae was still pending.
The present work i.e. Flora of Vietnam, volume 15 – Asclepiadaceae R. Br. is a taxonomic monograph, the output of the studies carried out for many years. To bring out this volume, specimens of the family Asclepiadaceae were collected in many regions of Vietnam with ecological observations. Specimens were stored at several herbaria such as Herbarium, Institute of Ecology and Biological Resources Hanoi (HN); Herbarium, Hanoi University of Science, National University Hanoi (HNU); Herbarium National Institute of Medicinal Materials, Hanoi (HNPM); Herbarium, Hanoi University of Pharmacy, Hanoi (HNPI); Herbarium, Institute of Tropical Biology, Ho Chi Minh city (HM, VNM); etc in Vietnam; and other countries like Kunming Botanical Institute, Academia Sinica, Yunnan, China (KUN); Botanical Institute, Faculty of Science, University of Tokyo, Hongo, Japan (TI); Herbarium of Kyoto University, Japan (KYO); Museum National d’Histoire Naturalle, Paris, France (P); Herbarium, Guangxi Institute of Botany Guangxb, Zhuangzu (IBK); Daejeon University, Republic of Korea; etc.
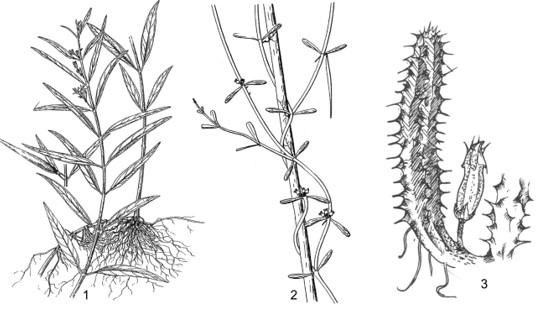
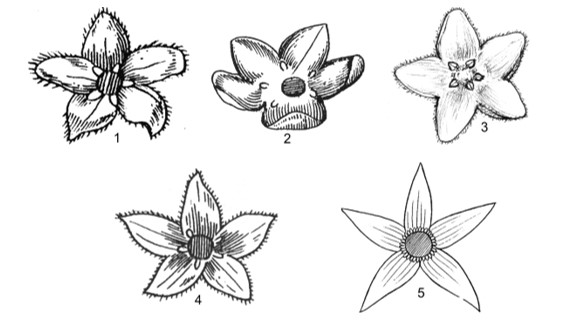
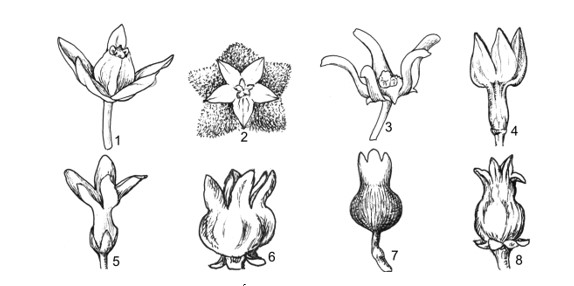
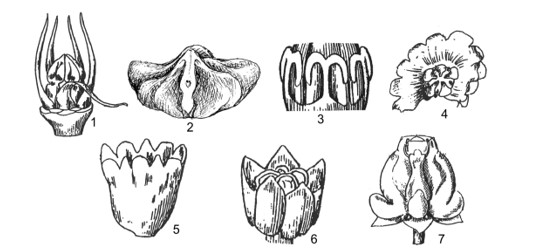
This work comprises 484 pages, presenting contents of morphological characters, and evaluates different taxonomic systems of classification. Keys to taxa, updated nomenclature, synonyms, morphological description, types, ecology, distribution, studied specimens, uses, remarks are provided. 125 figures and 120 tables of photos, indexes, references are provided.
Literatures, figures and photos from other sources are also used and properly acknowledged in this study.
Taxonomic system of Asclepiadaceae in Vietnam based on S. Liede & F. Albers (1994):
I. CHECKLIST OF THE TAXA IN THE
FAMILY ASCLEPIADACEAE IN VIETNAM
(3 subfam., 6 trib., 49 gen., 143 sp.; 1 gen., 13 sp., not information)
Subfam.1. PERIPLOCOIDEAE R. Br. ex Endl. (1 trib., 11 gen., 16 sp.).
Trib.1. PERIPLOCEAE (11 gen, 16 sp.)
1. TELECTADIUM Baill. (3 sp.)
1.1. Telectadium edule Baill. (fig. 11, photo 25)
1.2. Telectadium linearicarpum Pierre ex Cost. (fig. 12, photo 26)
1.3. Telectadium dongnaiense Pierre ex Cost. (fig. 13, photo 27)
2. HEMIDESMUS R. Br. (1 sp.)
2.1. Hemidesmus indicus (L.) R. Br. ex Schult. (fig. 14, photo 28)
3. CRYPTOLEPIS R. Br. (2 sp.)
3.1. Cryptolepis buchananii Roem. & Schult. (fig.15, photo 29)
3.2. Cryptolepis sinensis (Lour.) Merr. (fig. 16, photo 30)
4. ZYGOSTELMA Benth. (1 sp.)
4.1. Zygostelma benthamii Baill. (fig.17, photo 31)
5. GYMNANTHERA R. Br. (1 sp.)
5.1. Gymnanthera oblonga (Burm. f.) P. S. Green (fig.18, photo 32)
6. FINLAYSONIA Wall. (1 sp.)
6.1. Finlaysonia obovata Wall. (fig. 19, photo 33)
7. ATHEROLEPIS Hook. f. (1 sp.)
7.1. Atherolepis pierrei Cost. (fig. 20)
8. ATHERANDRA Decne. (1 sp.)
8.1. Atherandra acutifolia Decne. (fig. 21)
9. PERIPLOCA L. (1 sp.)
9.1. Periploca calophylla (Wight & Arn.) Falc. (fig. 22, photo 34)
10. STREPTOCAULON Wight & Arn. (3 sp.)
10.1. Streptocaulon juventas (Lour.) Merr. (fig. 23, photo 35)
10.2. Streptocaulon kleinii Wight & Arn. (fig. 24, photo 36)
10.3. Streptocaulon wallichii Wight (fig. 25)
11. MYRIOPTERON Griff. (1 sp.)
11.1. Myriopteron extensum (Wight & Arn.) K. Schum. (fig. 26, photo 37)
Subfam. 2. SECAMONOIDEAE Endl. (1 trib., 3 gen, 9 sp.; 2 sp. not information)
Trib.2. SECAMONEAE [3 gen., 9 sp. (2 sp. not information)]
12. TOXOCARPUS Wight & Arn. [7 sp. (2 sp. not information)]
12.1. Toxocarpus wightianus Hook. & Arn. (fig. 27, photo 38)
12.2. Toxocarpus spirei Cost. (fig. 28, photo 39)
12.3. Toxocarpus villosus (Blume) Decne. (fig. 29, photo 40)
12.4. Toxocarpus bonii Cost. (fig. 30)
12.5. Toxocarpus pierrei Cost. (fig. 31)
12.6. Toxocarpus gagnepainii Tsiang
12.7. Toxocarpus klossii S. Moore
13. GENIANTHUS Hook. f. (1 sp.)
13.1. Genianthus laurifolius (Roxb.) Hook. f. (fig. 32, photo 41)
14. SECAMONE R. Br. (1 sp.)
14.1. Secamone elliptica R. Br. (fig. 33, photo 42)
Subfam. 3. ASCLEPIADOIDEAE (4 trib., 34 gen., 117 sp. (1 gen., 10 sp. not information)]
Trib.3. FOCKEAE Kunze & al. (1 gen., 2 sp.)
15. FOCKEA Endl. (1 sp.)
15.1. Fockea glabra Decne. (fig. 34)
Trib.4. MARSDENIEAE Benth. [17 gen., 80 sp. (5 sp. not information)]
16. HOYA R. Br. [31 sp. (2 sp. not information)]
16.1. Hoya ignorata T. B. Tran, Rodda, Simonsson & Joongku Lee (photo 43)
16.2. Hoya multiflora Blume (fig. 35, photo 44)
16.3. Hoya lockii V. T. Pham & Aver. (photo 45)
16.4. Hoya chinghungensis (Tsiang & Li) M. G. Gilbert, P. T. Li & W. D. Stevens (fig. 36)
16.5. Hoya pubens Cost. (fig. 37)
16.6. Hoya macrophylla Blume (fig. 38, photo 46)
16.7. Hoya hanhiae V. T. Pham et Aver. (photo 47)
16.8. Hoya parasitica (Roxb.) Wall. ex Wight (fig. 39, photo 48)
16.9. Hoya balansae Cost. (fig. 40, photo 49)
16.10. Hoya nummularia Decne. ex Hook. f. (fig. 41)
16.11. Hoya minima Cost. (fig. 42)
16.12. Hoya pseudovalifolia Cost. (fig. 43)
16.13. Hoya griffithii Hook. f. (fig. 44, photo 50)
16.14. Hoya obovata Decne. (fig. 45, photo 51)
16.15. Hoya sapaensis T. B. Tran & Rodda (fig. 46, photo 52)
16.16. Hoya siamica Craib (photo 53)
16.17. Hoya thuathienhuensis T. B. Tran, Rodda & Simonsson (photo 54)
16.18. Hoya lyi H. Léveillé (photo 55)
16.19. Hoya esculenta (Rumph.) Tsiang (fig. 47, photo 56)
16.20. Hoya villosa Cost. (fig. 48, photo 57)
16.21. Hoya globulosa Hook. f.
16.22. Hoya oblongacutifolia Cost.
16.23. Hoya fungii Merr. (fig. 49)
16.24. Hoya fusca Wall. (fig. 50, photo 58)
16.25. Hoya tamdaoensis Rodda & T. B. Tran (photo 59)
16.26. Hoya graveolens Kerr (photo 60)
16.27. Hoya bonii Cost. (fig. 51, photo 61)
16.28. Hoya carnosa (L.) R. Br. (fig. 52, photo 62)
16.29. Hoya pubicalyx Merr. (photo 63)
16.30. Hoya cochinchinensis (Lour.) Roem & Schult
16.31. Hoya wallichiana Decne.
17. DISCHIDIA R. Br. [13 sp. (1 sp. not infomation)]
17.1. Dischidia major (Vahl) Merr. (fig. 53, photo 64)
17.2. Dischidia hirsuta (Blume) Decne. (fig. 54, photo 65)
17.3. Dischidia nummularia R. Br. (fig. 55, photo 66)
17.4. Dischidia bengalensis Colebr. (fig. 56, photo 67)
17.5. Dischidia tonkinensis Cost. (fig. 57, photo 68)
17.6. Dischidia imbricata (Blume) Steud. (fig. 58, photo 69)
17.7. Dischidia singularis Craib (fig. 59, photo 70)
17.8. Dischidia rimicola Kerr (fig. 60, photo 71)
17.9. Dischidia griffithii Griff. ex Hook. f. (fig. 61, photo 72)
17.10. Dischidia acuminata Cost. (fig. 62, photo 73)
17.11. Dischidia acutifolia Maingay ex Hook. f. (fig. 63, photo 74)
17.12. Dischidia dohtii Tran & Livsh. (fig. 64, photo 75)
17.13. Dischidia wallichii Wight
18. LYGISMA Hook. f. (1 sp.)
18.1. Lygisma inflexum (Cost.) Kerr (fig. 65)
19. SARCOLOBUS R. Br. (2 sp.)
19.1. Sarcolobus globosus Wall. (fig. 66)
19.2. Sarcolobus pierrei Cost. (fig. 67)
20. PSEUDOSARCOLOBUS Cost. (1 sp.)
20.1. Pseudosarcolobus villosus Cost. (fig. 68)
21. SPIRELLA Cost. (1 sp.)
21.1. Spirella robinsonii Cost. (fig. 69)
22. GYMNEMOPSIS Cost. (1 sp.)
22.1. Gymnemopsis pierrei Cost. (fig. 70)
23. HARMANDIELLA Cost. (1 sp.)
23.1. Harmandiella cordifolia Cost. (fig. 71)
24. GYMNEMA R. Br. [7 sp. (2 sp. not information)]
24.1. Gymnema inodorum (Lour.) Decne. (fig. 72, photo 76)
24.2. Gymnema yunnanense Tsiang (fig. 73, photo 77)
24.3. Gymnema sylvestre (Retz.) R. Br. ex Schult. (fig. 74, photo 78)
24.4. Gymnema latifolium Wall. ex Wight (fig. 75, photo 79)
24.5. Gymnema reticulatum (Moon) Alston
24.6. Gymnema foetidum Tsiang
24.5. Gymnema griffithii Craib
25. DISCHIDANTHUS Tsiang (1 sp.)
25.1. Dischidanthus urceolatus (Decne.) Tsiang (fig. 76, photo 80)
26. MARSDENIA R. Br. (7 sp.)
26.1. Marsdenia tenacissima (Roxb.) Moon (fig. 77, photo 81)
26.2. Marsdenia hainanensis Tsiang (fig. 78, photo 82)
26.3. Marsdenia koi Tsiang (fig. 79)
26.4. Marsdenia balansae Cost. (photo 83)
26.5. Marsdenia glabra Cost. (fig. 80)
26.6. Marsdenia tinctoria R. Br. (fig. 81, photo 84)
26.7. Marsdenia tonkinensis Cost. (photo 85)
27. DREGEA E. Mey. (2 sp.)
27.1. Dregea volubilis (L. f.) Benth. & Hook. (fig. 82, photo 86)
27.2. Dregea cuneifolia Tsiang & P. T. Li (fig. 83)
28. COSMOSTIGMA Wight (1 sp.)
28.1. Cosmostigma racemosum (Roxb.) Wight (fig. 84, photo 87)
29. TELOSMA Coville (2 sp.)
29.1. Telosma cordata (Burm. f.) Merr. (fig. 85, photo 88)
29.2. Telosma procumbens (Blanco) Merr. (fig. 86, photo 89)
30. CAMPESTIGMA Pierre ex Cost. (1 sp.)
30.1. Campestigma purpurea Pierre ex Cost. (fig. 87, photo 90)
31. PENTASACME Wall. ex Wight (1 sp.)
31.1. Pentasacme caudatum Wall. ex Wight (fig. 88, photo 91)
32. HETEROSTEMMA Wight & Arn. (7 sp.)
32.1. Heterostemma xuansonense T. B. Tran & Joo-Hwan Kim (fig. 89, photo 92)
32.2. Heterostemma oblongifolium Cost. (fig. 90)
32.3. Heterostemma grandiflorum Cost. (fig. 91, photo 93)
32.4. Heterostemma brownii Hayata (fig. 92, photo 94)
32.5. Heterostemma piperifolium King & Gamble (photo 95)
32.6. Heterostemma acuminatum Decne. (fig. 93)
32.7. Heterostemma suberosum Cost. (fig. 94, photo 96)
Trib.5. CEROPEGIEAE Orb. [5 gen., 9 sp. (2 sp. not information available)]
33. CEROPEGIA L. [4 sp. (2 sp. not information)]
33.1. Ceropegia candelabrum L. (fig. 95, photo 97)
33.2. Ceropegia driophila C. K. Schneid. (fig. 96, photo 98)
33.3. Ceropegia obtusa Lour.
33.4. Ceropegia cordata Lour.
34. TAVARESIA Welw. (1 sp.)
34.1. Tavaresia barklyi N. E. Br. (fig. 97, photo 99)
35. HUERNIA R. Br. (1 sp.)
35.1. Huernia transvaalensis Stent (fig. 98, photo 100)
36. STAPELIA L. (2 sp.)
36.1. Stapelia grandiflora Mass. (fig. 99, photo 101)
36.2. Stapelia puchella Mass. (fig. 100)
37. ECHIDNOPSIS Hook. f. (1 sp.)
37.1. Echidnopsis tessellata K. Schum. (fig. 101, photo 102)
Trib. 6. ASCLEPIADEAE [11 gen., 27 sp. (3 sp. not information)]
38. SARCOSTEMMA R. Br. (1 sp.)
38.1. Sarcostemma acidum (Roxb.) Voigt (fig. 102, photo 103)
39. OXYSTELMA R. Br. (1 sp.)
39.1. Oxystelma esculentum (L. f) R. Br. ex Schult. (fig. 103, photo 104)
40. RAPHISTEMMA Wall. (2 sp.)
40.1. Raphistemma hooperianum (Blume) Decne. (fig. 104, photo 105)
40.2. Raphistemma pulchellum (Roxb.) Wall. (fig. 105, photo 106)
41. CYNANCHUM L. [6 sp. (2 sp. not information)]
41.1. Cynanchum corymbosum Wight (fig. 106, photo 107)
41.2. Cynanchum officinale (Hemsl.) Tsiang & Zhang (fig. 107, photo 108)
41.3. Cynanchum utriculosum Cost. (fig. 108, photo 109)
41.4. Cynanchum stauntonii (Decne.) Hand.-Mazz. (fig. 109)
41.5. Cynanchum medium (Decne.) K. Schum.
41.6. Cynanchum verticillatum Hemsl.
42. PENTATROPIS R. Br. ex Wight & Arn. (2 sp.)
42.1. Pentatropis pierrei Cost. (fig. 110, photo 110)
42.2. Pentatropis capensis (L. f.) Bull. (fig. 111)
43. MERRILLANTHUS Chun & Tsiang (1 sp.)
43.1. Merrillanthus hainanensis Chun & Tsiang (fig. 112, photo 111)
44. TYLOPHORA R. Br. [9 sp. (1 sp. not information)]
44.1. Tylophora ovata (Lindl.) Hook. & Steud. (fig. 113, photo 112)
44.2. Tylophora flexuosa R. Br. (fig. 114, photo 113)
44.3. Tylophora koi Merr. (fig. 115, photo 114)
44.4. Tylophora kerrii Craib (fig. 116)
44.5. Tylophora indica (Burm. f.) Merr. (fig. 117, photo 115)
46.6. Tylophora harmandii Cost. (fig. 118)
44.7. Tylophora glabra Cost. (fig. 119)
44.8. Tylophora tengii Tsiang (fig. 120)
44.9. Tylophora dalatensis S. Moore
45. ASCLEPIAS L. (1 sp.)
45.1. Asclepias curassavica L. (fig. 121, photo 116)
46. VINCETOXICOPSIS Cost. (1 sp.)
46.1. Vincetoxicopsis harmandii Cost. (fig. 122, photo 117)
47. GOMPHOCARPUS R. Br. (1 sp.)
47.1. Gomphocarpus fruticosus (L.) Ait. f. (fig. 123, photo 118)
48. CALOTROPIS R. Br. (2 sp.)
48.1. Calotropis gigantea (L.) Dryand. ex Ait. f. (fig. 124, photo 119)
48.2. Calotropis procera (Ait.) Dryand. ex Ait. f. (fig. 125, photo 120)
49. CRYPTOSTEGIA R. Br. [Not information]
49.1. Cryptostegia elegans Vahl
II. KEY TO THE GENERA OF ASCLEPIADACEAE IN VIETNAM
| 1A. |
Anthers 4-celled, pollen tetrads or pollen united into pollinia without waxy. |
|
| 2A. |
Pollen tetrads or pollen united into pollinia on a spatulate translator with a viscid basal disc. |
(Subfam.1. PERIPLOCOIDEAE) (Trib.1. Periploceae) |
| 3A. |
Erect shrubs. |
1. TELECTADIUM |
| 3B. |
Climbing or prostrate plants. |
|
| 4A. |
Corona inserted on corolla tube. |
|
| 5A. |
Corolla lobes valvate. |
2. HEMIDESMUS |
| 5B. |
Corolla lobes overlapping. |
|
| 6A. |
Corona lobes are not divided. |
3. CRYPTOLEPIS |
| 6B. |
Corona lobes are divided. |
4. ZYGOSTELMA |
| 4B. |
Corona separate from corolla. |
|
| 7A. |
The middle of filaments without glands. |
|
| 8A. |
Corona lobes ovate. |
5. GYMNANTHERA |
| 8B. |
Corona lobes thread-shaped. |
6. FINLAYSONIA |
| 7B. |
The middle of filaments with glands. |
|
| 9A. |
Inside of calyx without basal glands. |
|
| 10A. |
Corona lobes short, triangular. |
7. ATHEROLEPIS |
| 10B. |
Corona lobes long, thread-shaped. |
8. ATHERANDRA |
| 9B. |
Inside of calyx with basal glands. |
|
| 11A. |
Anther hair. Corona lobes 3-fid, central segment threadlike, inflexed; lateral segments short, broad, flaplike. |
9. PERIPLOCA |
| 11B. |
Anther glabrous. Corona lobes unlike above characters. |
|
| 12A. |
Follicles without wings, pubescent. |
10. STREPTOCAULON |
| 12B. |
Follicle with wings, glabrous. |
11. MYRIOPTERON |
| 2B. |
Pollens aggregate into pollinia in translator clip-shaped, translator with corpusculum. |
(Subfam.2. SECAMONOIDEAE)......(Trib.2. Secamoneae). |
| 13A. |
Apex of anther without membranous appendage. Flower large (diam.> 5 mm). |
12. TOXOCARPUS |
| 13B. |
Apex of anther with membranous appendage. Flower small (diam. < 5 mm). |
|
| 14A. |
Inside of corolla lobes hair, corona lobes triangular. |
13. GENIANTHUS |
| 14B. |
Inside of corolla lobes hair, corona lobes sickle-shaped. |
14. SECAMONE |
| 1B. |
Anther 2-celled, pollens aggregate into pollinia, outside of pollinium walled. |
(Subfam.3. ASCLEPIADOIDEAE) |
| 15A. |
Translator without caudicles |
(Trib.3. Fockeae)........15. FOCKEA |
| 15B. |
Translator with caudicles. |
|
| 16A. |
Pollinium erect; apex of ovary not narrow into style-shaped. |
|
| 17A. |
Apex of anther with appendage. |
(Trib.4. Marsdenieae). |
| 18A. |
Apex and inside of pollinium without crests. |
|
| 19A. |
Plants always growing on trees or rock surfaces, often using adventitious roots; Leaves fleshy. |
|
| 20A. |
Corolla rotate, corona fleshy. |
16. HOYA |
| 20B. |
Corolla urceolate, corona non-fleshy. |
17. DISCHIDIA |
| 19B. |
Climbing, non-epithytic, without adventitious roots on stem. Leaves non-fleshy. |
|
| 21A. |
Corolla lobes folded in flower bud. |
18. LYGISMA |
| 21B. |
Corolla lobes non-folded in flower bud. |
|
| 22A. |
Without corona. |
|
| 23A. |
Gynostegium with short stalk. |
19. SARCOLOBUS |
| 23B. |
Gynostegium without a short stalk. |
|
| 24A. |
Flower bud globose. Calyx obtuse, hair. |
20. PSEUDOSARCOLOBUS |
| 24B. |
Flower hat-shaped. Calyx acute, glabrous. |
21. SPIRELLA |
| 22B. |
With corona. |
|
| 25A. |
Corona simple. |
|
| 26A. |
Corona inserted on corolla. |
|
| 27A. |
Stigma non-swelled. |
|
| 28A. |
Corpusculum swelled. |
22. GYMNEMOPSIS |
| 28B. |
Corpusculum sunken. |
23. HARMANDIELLA |
| 27B. |
Stigma swelled. |
24. GYMNEMA |
| 26B. |
Corona non-inserted on corolla. |
|
| 29A. |
Corolla urceolate. |
|
| 30A. |
Inside of corolla lobes with the swelled edge. |
25. DISCHIDANTHUS |
| 30B. |
Inside of corolla lobes without the swelled edge. |
26. MARSDENIA |
| 29B. |
Corolla rotate or trumpet-shaped. |
|
| 31A. |
Corolla rotate, united parts non tube-shaped. |
|
| 32A. |
Corona fleshy. |
27. DREGEA |
| 32B. |
Corona non fleshy. |
28. COSMOSTIGMA |
| 31B. |
Corolla trumpet-shaped, united parts tube-shaped. |
29. TELOSMA |
| 25B. |
Corona double. |
30. CAMPESTIGMA |
| 18B. |
Apex and inside of pollinium with crests. |
|
| 33A. |
Plants erect. |
31. PENTASACME |
| 33B. |
Plants climbing or prostrate. |
32. HETEROSTEMMA |
| 17B. |
Apex of anther without appendage. |
(Trib.5. Ceropegieae). |
| 34A. |
Stem non-angular. Flower non-fleshy. |
33. CEROPEGIA |
| 34B. |
Stem angular (like Cactus stem). Flower fleshy. |
|
| 35A. |
Inside of corolla with hard hair. |
|
| 36A. |
Pollinium D-shaped, length of corpusculum > 2 times of width of corpusculum. |
34. TAVARESIA |
| 36B. |
Pollinium elliptic, length of corpusculum < 2 times of width of corpusculum. |
35. HUERNIA |
| 35B. |
Inside of corolla without hard hair. |
|
| 37A. |
Inside of corolla with soft hair, pollinium D-shaped. |
36. STAPELIA |
| 37B. |
Inside of corolla glabrous, pollinium elliptic. |
37. ECHIDNOPSIS |
| 16B. |
Pollinium pendulous; pollinium erect; apex of ovary narrow into style-shaped. |
(Trib.6. Asclepiadeae) |
| 38A. |
Corona double. |
|
| 39A. |
Leaves reduced. |
38. SARCOSTEMMA |
| 39B. |
Leaves developed. |
39. OXYSTELMA |
| 38B. |
Corona simple. |
|
| 40A. |
Corona lobes united. |
|
| 41A. |
Flower large (Diam. > 1.5 cm). |
40. RAPHISTEMMA |
| 41B. |
Flower small (Diam. < 8 mm). |
|
| 42A. |
Corona cup-shaped. |
41. CYNANCHUM |
| 42B. |
Corona non cup-shaped. |
42. PENTATROPIS |
| 40B. |
Corona lobes free. |
|
| 43A. |
Climbing plants. |
|
| 44A. |
Axis of inflorescence fleshy. Corolla lobe 4 mm. Skin of fruit thick. |
43. MERRILLANTHUS |
| 44B |
Axis of inflorescence slender. Corolla lobes < 3 mm. Skin of fruit thin. |
44. TYLOPHORA |
| 43B. |
Erect plants. |
|
| 45A. |
Corona lobes with appendage horn-shaped. |
45. ASCLEPIAS |
| 45B. |
Corona lobes without appendage horn-shaped. |
|
| 46A. |
Corona lobe without basal spur. |
|
| 47A. |
Fruit without spine . |
46. VINCETOXICOPSIS |
| 47B. |
Fruit with spine. |
47. GOMPHOCARPUS |
| 46B. |
Corona lobe with basal spur. |
48. CALOTROPIS |
| |
Not information |
49. CRYPTOSTEGI |
III. KEY TO THE SPECIES OF ASCLEPIADACEAE IN VIETNAM
3.1. KEY TO THE SPECIES OF TELECTADIUM IN VIETNAM
| 1A. |
Translator ≥ 2 mm, tube of corolla is not divided into two parts. |
|
| 2A. |
Fruit ellptic |
1. T. edule |
| 2B. |
Fruit linear |
2. T. linearicarpum |
| 1B. |
Translator 1.1-1.4 mm, tube of corolla is divided into two parts (swelled part globose, above part cylindrical). |
3. T. dongnaiense |
3.2. KEY TO THE SPECIES OF HEMIDESMUS IN VIETNAM
| Climbing or prostrate plants; corona inserted on corolla tube; corolla lobes valvate |
1. H. indicus |
3.3. KEY TO THE SPECIES OF CRYPTOLEPIS IN VIETNAM
| 1A. |
Lateral veins 25-30 pairs, calyx glabrous. |
1. C. buchananii |
| 1B. |
Lateral veins 5-9 pairs, calyx pubescent |
2. C. sinensis |
3.4. KEY TO THE SPECIES OF ZYGOSTELMA IN VIETNAM
| Corolla lobes overlapping; corona lobes are divided. |
1. Z. benthamii |
3.5. KEY TO THE SPECIES OF GYMNANTHERA IN VIETNAM
| The middle of filaments without glands; corona lobes ovate |
1. G. oblonga |
3.6. KEY TO THE SPECIES OF FINLAYSONIA IN VIETNAM
| The middle of filaments without glands; corona lobes thread-shaped |
1. F. obovata |
3.7. KEY TO THE SPECIES OF ATHEROLEPIS IN VIETNAM
| Inside of calyx without basal glands; corona lobes short, triangular |
1. A. pierrei |
3.8. KEY TO THE SPECIES OF ATHERANDRA IN VIETNAM
| Inside of calyx without basal glands; corona lobes long, thread-shaped |
1. A. acutifolia |
3.9. KEY TO THE SPECIES OF PERIPLOCA IN VIETNAM
| Anther hair. Corona lobes 3-fid, central segment threadlike, inflexed; lateral segment short, broad, flaplike |
1. P. calophylla |
3.10. KEY TO THE SPECIES OF STREPTOCAULON IN VIETNAM
| 1A. |
Pedicell hair. |
|
| 2A. |
Lateral veins 14-20 pairs. Translator ca. 350 mm long |
1. S. juventas |
| 2B. |
Lateral veins <13 pairs. Translator ca. 540 mm long |
2. S. kleinii |
| 1B. |
Pedicell glabrous |
3. S. wallichii |
3.11. KEY TO THE SPECIES OF MYRIOPTERON IN VIETNAM
| Anther glabrous. Corona lobes unlike obove characters. Follicle with wings, glabrous. |
1. M. extensum |
3.12. KEY TO THE SPECIES OF TOXOCARPUS IN VIETNAM
| 1A. |
Stem haired. |
|
| 2A. |
Style swell at apex |
1. T. wightianus |
| 2B. |
Style not swell at apex. |
|
| 3A. |
Apex of style is divided into two lobes. |
|
| 4A. |
Style short; flower bud < 8mm |
2. T. spirei |
| 4B. |
Style long; flower bud > 8 mm |
3. T. villosus |
| 3B. |
Apex of style is not divided |
4. T. bonii |
| 1B. |
Stem glabrous |
5. T. pierrei |
| |
Not information |
6. T. gagnepainii |
| |
|
7. T. klosii |
3.13. KEY TO THE SPECIES OF GENIANTHUS IN VIETNAM
| Apex of anther with membrannous appendge. Flower small. Inside of corolla bobes hair, corona lobes triangular |
1. G. laurifolius |
3.14. KEY TO THE SPECIES OF SECAMONE IN VIETNAM
| Apex of anther with membrannous appendge. Flower small. Inside of corolla bobes hair, corona lobes stickle –shaped |
1. S. elliptica |
3.15. KEY TO THE SPECIES OF FOCKEA IN VIETNAM
| Anther 2-celled, pollens aggregate into pollina, outsite of pollinium with wall. Transtor with caudicles |
1. F. glabra |
3.16. KEY TO THE SPECIES OF HOYA IN VIETNAM
| 1A. |
Erect plants. |
|
| 2A. |
Flower < 4 mm diam. |
1. H. ignorata |
| 2B. |
Flower > 6 mm diam. |
|
| 3A. |
Corona distinctly stalked. |
|
| 3B. |
Corona sessile. |
|
| 4A. |
Leave > 10 cm long |
3. H. lockii |
| 4B. |
Leave < 3 cm long |
4. H. chinghungensis |
| 1B. |
Twining pendulous or rambling. |
|
| 6A. |
Pollinium 0.4 mm long |
5. H. pubens |
| 6B. |
Pollinium > 0.5 mm long. |
|
| 7A. |
Flower ca. 18 mm diam |
6. H. macrophylla |
| 7B. |
Flower < 15 mm diam. |
|
| 8A. |
Inner corona purple |
7. H. hanhiae |
| 8B. |
Inner corona otherwise. |
|
| 9A. |
Corolla lobes < 4 mm long; corona lobes < 3 mm long; pollinium 0.5 mm long, corpusculum 0.25 mm long |
8. H. parasitica |
| 9B. |
Corolla lobes > 4.5 mm long; corona lobes 4 mm long; pollinium 0.7 mm long, corpusculum 0.3 mm long |
9. H. balansae |
| 5B. |
Leave pinnately veined, or veins obscure. |
|
| 10A. |
Leave < 2 cm long. |
10. H. nummularia |
| 10B. |
Leave ≥ 2 cm long. |
|
| 11A. |
Leave ≤ 1.5 cm wide |
11. H. minima |
| 11B. |
Leave > 2 cm wide. |
|
| 12A. |
Pedicel < 5 mm long |
12. H. pseudovalifolia |
| 12B. |
Pedicel ≥ 7 mm long. |
|
| 13A. |
Outer angle of corona lobes rounded or obtuse. |
|
| 14A. |
Flower ca. 3 cm diam |
13. H. griffithii |
| 14B. |
Flower ca. < 2.5 cm diam. |
|
| 15A. |
Corona red throughout |
14. H. obovata |
| 15B. |
Corona otherwise. |
|
| 16A. |
Inflorescence umbel, few-flowered (less than 15). |
|
| 17A. |
Peduncle < 2 cm; corolla 17-20 mm diam.; pollinium 0.95 mm long, 0.41 mm wide |
15. H. sapaensis |
| 17B. |
Peduncle > 4 cm; corolla 8-9.4 mm diam.; pollinium < 0.8 mm long, < 0.3 mm wide |
16. H. siamica |
| 16B. |
Inflorescence umbel, many-flowered (more than 20). |
|
| 18A. |
Apex of leave with cuspidate caudate. |
17. H. thuathienhuensis |
| 18B. |
Apex of leave without cuspidate caudate. |
|
| 19A. |
Peduncle up to 10 cm long |
18. H. lyi |
| 19B. |
Maximum peduncle 8 cm long. |
|
| 20A. |
Fruit < 18 cm long . |
|
| 21A. |
Pollinium 0.54 mm long, corpusculum ca. 0.2 mm long |
19. H. esculenta |
| 21B. |
Pollinium 0.9 mm long, corpusculum ca. 0.4 mm long |
20. H. villosa |
| 20B. |
Fruit > 20 cm long |
21. H. globulosa |
| 13B. |
Outer angle of corona lobe cuspidate. |
|
| 22A. |
Leave non-fleshy |
22. H. oblongacutifolia |
| 22B. |
Leave fleshy. |
|
| 23A. |
Pollinium 0.4 mm long |
23. H. fungii |
| 23B. |
Pollinium >0.6 mm long |
|
| 24A. |
Lateral veins ≥ 10 pairs. |
24. H. fusca |
| 24B. |
Lateral veins < 8 pairs. |
|
| 25A. |
Inner corolla lobes white. |
|
| 26A. |
Corpusculum 0.3-0.4 mm long |
|
| 27A. |
Apex of leave with long caudate, lobes of corona orange at top (center of flower) |
25. H. tamdaoensis |
| 27B. |
Apex of leave with short caudate, lobes of corona purple at top (center of flower) |
26. H. graveolens |
| 26B. |
Corpusculum <0.25 mm long. |
|
| 28A. |
Corpusculum 0.125 mm long. Inner corona purple. |
27. H. bonii |
| 28B. |
Corpusculum 0.2 mm long. Inner corona red |
28. H. carnosa |
| 25B. |
Inner corolla lobes red-purple |
29. H. pubicalyx |
| |
Not information . |
30. H. cochinchinensis |
| |
|
31. H. wallichiana |
3.17. KEY TO THE SPECIES OF DISCHIDIA IN VIETNAM
| 1A. |
Staminal corona lobes anchor-shaped or sagittate. |
|
| 2A. |
Corona lobes anchor-shaped. |
|
| 3A. |
Root-filled pitcher leaves present. |
1. D. major |
| 3B. |
Root-filled pitcher leaves absent. |
|
| 4A. |
Corolla 6-9 mm long, with 2 rings of hais: the ring of hair at throat, other ring below corolla lobes. |
2. D. hirsuta |
| 4B. |
Corolla ≤ 5 mm, without 2 rings of hairs. |
|
| 5A. |
Corolla lobes glabrous |
3. D. nummularia |
| 5B. |
Corolla lobes haired. |
|
| 6A. |
Leaves linear |
4. D. bengalensis |
| 6B. |
Leaves otherwise |
5. D. tonkinensis |
| 2B. |
Corona lobes sagittate. |
|
| 7A. |
Corolla lobes haired. |
|
| 8A. |
Leaves reniform. Peduncle ≥ 2 cm |
6. D. imbricata |
| 8B. |
Leaves otherwise. Peduncle ≤ 1 cm. |
|
| 9A. |
Corolla 4.8-5 mm long, without longitudinal nerves inside corolla, corona base glabrous. |
7. D. singularis |
| 9B. |
Corolla 3.5-4 mm long, with longitudinal nerves inside corolla, corona base haired |
8. D. rimicola |
| 7B. |
Corolla lobes glabrous. |
|
| 10A. |
Leaves obovate, oblanceolate. Caudicle shoter than corpusculum |
9. D. griffithii |
| 10B. |
Leaves otherwise. Caudicle longer than corpusculum. |
|
| 11A. |
Corolla tube > 2.5 mm |
10. D. acuminata |
| 11B. |
Corolla tube ≤ 2.5 mm |
11. D. acutifolia |
| 1B. |
Staminal corona absent |
12. D. dohtii |
| |
Not information |
13. D. wallichii |
3.18. KEY TO THE SPECIES OF LYGISMA IN VIETNAM
| Climbing non-epithytic, without adventitious roots on stem. Leaf nonfleshy. Corolla lobes folded in flower bud |
1. L. inflexum |
3.19. KEY TO THE SPECIES OF SARCOLOBUS IN VIETNAM
| 1A. |
Pedicel < 1.5 cm; base obtuse to shallowly cordate |
1. S. globosus |
| 1B. |
Pedicel ≥ 1.5 cm; base cordate |
2. S. pierrei |
3.20. KEY TO THE SPECIES OF PSEUDOSARCOLOBUS IN VIETNAM
| Gynostegium without short stalk. Flower bud globose. Calyx obtuse, hair |
1. P. villosus |
3.21. KEY TO THE SPECIES OF SPIRELLA IN VIETNAM
| Gynostegium without short stalk. Flower hat-shaped. Calyx acute, glabrous |
1. S. robinsonii |
3.22. KEY TO THE SPECIES OF GYMNEMOPSIS IN VIETNAM
| Corona inserted on corolla. Stigma non-swelled. Corpusculum swelled |
1. G. pierrei |
3.23. KEY TO THE SPECIES OF HARMANDIELLA IN VIETNAM
| Corona inserted on corolla. Stigma non-swelled. Corpusculum swelled |
1. H. cordifolia |
3.24. KEY TO THE SPECIES OF GYMNEMA IN VIETNAM
| 1A. |
Follicle > 12 cm long |
1. G. inodorum |
| 1B. |
Follicle < 10 cm long. |
|
| 2A. |
Coma < 2.6 cm |
2. G. yunnanense |
| 2B. |
Coma ≥ 3 cm. |
|
| 3A. |
Seed < 0.9 cm long |
3. G. sylvestre |
| 3B. |
Seed > 1 cm long. |
|
| 4A. |
Follicle < 6 cm long; coma < 3.5 mm |
4. G. latifolium |
| 4B. |
Follicle > 7 cm; coma > 3.5 mm. |
5. G. reticulatum |
| 3B. |
Not information |
6. G. foetidum |
| |
|
6. G. foetidum |
| |
|
7. G. griffithii |
3.25. KEY TO THE SPECIES OF DISCHIDANTHUS IN VIETNAM
| Corona urceolate. Inside of corolla lobes with the swelled edge |
1. D. urceolatus |
3.26. KEY TO THE SPECIES OF MARSDENIA IN VIETNAM
| 1A. |
Inflorescence branched. |
|
| 2A. |
Outside of corolla haired; corona apex at a higher position of corolla tube, apex with 2(4) teeth |
1. M. tenacissima |
| 2B. |
Outside of corolla glabrous; corona apex below the position of corolla tube, apex without teeth |
2. M. hainanensis |
| 3A. |
Peduncle ≥7 cm |
3. M. koi |
| 3B. |
Peduncle ≤ 5 cm. |
|
| 4A. |
Axis of inflorescence twisted |
4. M. balansae |
| 4B. |
Axis of inflorescence straight. |
5. G. reticulatum |
| 5A. |
Corolla tube ca. 5 mm long |
5. M. glabra |
| 5B. |
Corolla tube < 4 mm. |
6. G. foetidum |
| 6A. |
Pedicel 3-5 mm long |
6. M. tinctoria |
| 6B. |
Pedicel 1 mm long |
7. M. tonkinensis |
3.27. KEY TO THE SPECIES OF DREGEA IN VIETNAM
1A. Leave blade broade ovate-cordate. Pollinium < 0.5 mm long. Ovaries haired ...........
................................................................................................................. 1. D. volubilis
1B. Leave blade oblongate. Pollinium 0.6 mm long. Ovaries glabrous .. 2. D. cuneifolia
3.28. KEY TO THE SPECIES OF COSMOSTIGMA IN VIETNAM
Corolla rotate, united parts non tube-shaped. Corolla non fleshy ......... 1. C. racemosum
3.29. KEY TO THE SPECIES OF TELOSMA IN VIETNAM
1A. Leave deeply heart-shaped .................................................................... 1. T. cordata
1B. Leave otherwise.............................................................................. 2. T. procumbens
3.30. KEY TO THE SPECIES OF CAMPESTIGMA IN VIETNAM
Corolla lobes non-folded in flower bud. Corona double .......................... 1. C. purpurea
3.31. KEY TO THE SPECIES OF PENTASACME IN VIETNAM
Apex and inside of pollinium with crests. Plants erect. ............................ 1. P. caudatum
3.32. KEY TO THE SPECIES OF HETEROSTEMMA IN VIETNAM
1A. Stem without edge.
2A. Peduncle absent......................................................................... 1. H. xuansonense
2B. Peduncle present
3A. Peduncle ≤ 2 mm.................................................................. 2. H. oblongifolium
3B. Peduncle > 4 mm.
4A. Fruit with 2 follicles.
5A. Fruit with 2 follicles > 60o................................................ 3. H. grandiflorum
5B. Fruit with 2 follicles < 45o.
6A. Corolla 4-5 mm long................................................................ 4. H. brownii
6B. Corolla 1 cm long............................................................. 5. H. piperifolium
4B. Fruit with 1 follicle................................................................... 6. H. acuminata
1B. Stem with edge.................................................................................. 7. H. suberosum
3.33. KEY TO THE SPECIES OF CEROPEGIA IN VIETNAM
1A. Corolla tube white. Calyx lobes 4 mm long................................. 1. C. candelabrum
1B. Corolla tube red-brown. Calyx lobes 1,3-3 mm long.......................... 2. C. driophila
Not information .............................................................................................. 3. C. obtusa
............................................................................................. 4. C. cordata
3.34. KEY TO THE SPECIES OF TAVARESIA IN VIETNAM
Stem angular (like Cactus stem). Flower fleshy. Inside of corolla with hard hair. Pollinium D-shaped, length of corpusculum > 2 times of width..................................................................... 1. T. barklyi
3.35. KEY TO THE SPECIES OF HUERNIA IN VIETNAM
Stem angular (like cactus stem). Flower fleshy. Inside of corolla with hard hair. Pollinium elliptic, length of corpusculum < 2 times of width of corpusculum ...............................................................
............................................................................................................ 1. H. transvaalensis
3.36. KEY TO THE SPECIES OF STAPELIA IN VIETNAM
1A. Corolla inside pubescence................................................................ 1. S. grandiflora
1B. Corolla inside glabrous.......................................................................... 2. S. puchella
3.37. KEY TO THE SPECIES OF ECHIDNOPSIS IN VIETNAM
Stem angular (like Cactus stem). Flower fleshy. Inside of corolla without hard hair, glabrous. Pollinium elliptic..................................................................................................................... 1. E. tessellata
3.38. KEY TO THE SPECIES OF SARCOSTEMMA IN VIETNAM
Pollinium pendulous, erect; apex of ovary narrow into style-shaped. Corona double. Leaves reduced 1. S. acidum
3.39. KEY TO THE SPECIES OF OXYSTELMA IN VIETNAM
Pollinium pendulous, erect; apex of ovary narrow into style-shaped. Corona double. Leaves developed 1. O. esculentum
3.40. KEY TO THE SPECIES OF RAPHISTEMMA IN VIETNAM
1A. Length of corolla lobes ≥ length of corolla tube ......................... 1. R. hooperianum
1B. Length of corolla lobes < length of corolla tube............................... 2. R. pulchellum
3.41. KEY TO THE SPECIES OF CYNANCHUM IN VIETNAM
1A. Liana.
2A. Peduncle, pedicel haired. Fruit > 7 cm long. Five times the length of caudicle
> length of corpusculum.
3A. Corolla glabrous. Fruit haired; coma 3-4 cm long .................. 1. C. corymbosum
3B. Corolla inside haired. Fruit glabrous; coma 2 cm long ..........................................
.......................................................................................................... 2. C. officinale
2B. Peduncle, pedicel glabrous. Fruit < 7 cm. Five times the length of caudicle
< length of corpusculum................................................................ 3. C. utriculosum
1B. Plant erect .......................................................................................... 4. C. stauntonii
Not information . .......................................................................................... 5. C. medium
.................................................................................. 6. C. verticillatum
3.42. KEY TO THE SPECIES OF PENTATROPIS IN VIETNAM
1A. Leave glabrous.......................................................................................... 1. P. pierrei
1B. Leave haired .......................................................................................... 2. P. capensis
3.43. KEY TO THE SPECIES OF MERRILLANTHUS IN VIETNAM
Corona lobes free. Climbing plant. Axis of inflorescence fleshy. Corolla lobe 4 mm. Skin of fruit thick 1. M. hainanensis
3.44. KEY TO THE SPECIES OF TYLOPHORA IN VIETNAM
1A. Pubescence around stem ............................................................................ 1. T. ovata
1B. Stem with 1-2 rows of pubesscence.
2A. Lateral veins obscure........................................................................... 2. T. flexuosa
2B. Lateral veins obvious.
3A. Length of inflorescence > length of leave................................................. 3. T. koi
3B. Length of inflorescence < length of leave.
4A. Leave linear or lanceolate .................................................................. 4. T. kerrii
4B. Leave otherwise.
5A. Peduncle haired................................................................................ 5. T. indica
5B. Peduncle glabrous.
6A. Leave > 4 cm long.
7A. Leave membrane, ovate-triangular ..................................... 6. T. harmandii
7B. Leave subfleshy, oval-oblong or lanceolate-oblong .................. 7. T. glabra
6B. Leave ≤ 4 cm long.......................................................................... 8. T. tengii
Not information ......................................................................................... 9. T. dalatensis
3.45. KEY TO THE SPECIES OF ASCLEPIAS IN VIETNAM
Erect plants. Corona lobes free. Erect plants. Corona lobe with appendage horn-shaped .
................................................................................................................ 1. A. curassavica
3.46. KEY TO THE SPECIES OF VINCETOXICOPSIS IN VIETNAM
Erect plants. Corona lobes without appendage horn-shaped, without basal spur. Fruit without spine 1. V. harmandii
3.47. KEY TO THE SPECIES OF GOMPHOCARPUS IN VIETNAM
Erect plants. Corona lobes without appendage horn-shaped, without basal spur. Fruit with spine 1. G. fruticosus
3.48. KEY TO THE SPECIES OF CALOTROPIS IN VIETNAM
1A. Corona white ....................................................................................... .1. C. gigantea
1B. Corona violet........................................................................................ ..2. C. procera
3.49. GENUS CRYPTOSTEGIA IN VIETNAM
Not information ........................................................................................... ..1. C. elegans








_254903291020251.jpg)






