ABSTRACT
FLORA OF VIETNAM 16
ARACEAE Juss. 1789
(4 subfamilies, 14 tribes, 24 genera, 138 species, 3 varieties, 1 forma)
INTRODUCTION
The family Araceae Juss. includes 106 genera and 2823 species, distributed in all continents, mainly in Asia and south America, mostly adapted to wet tropical weather. In Vietnam 24 genera, 138 species, 3 varieties and 1 forma have been recorded, widely distributed from north to south, occurring at the elevations ranging from sea level to high mountain areas. This family is important because of the presence of many economically important species, having medicinal and ornamental value.
The history of monographic works on the family Araceae dates back to 17th century. The pioneer work on Araceae was carried out by A. L. Jussieu (1789). Later H. W. Schott (1860), A. Engler (1876-1920), A. Engler & K. Krause (1908, 1920) also contributed to the taxonomy of Araceae. In Asian regions, J. D. Hooker (1893), M. Sivadasan (1982), M. Hotta (1970), S. Y. Hu (1968), Li Heng (1987-1992), D. Sriboonma (1994), P. C. Boyce (1996, 1998, 1999), W. L. A. Hetterscheid (1994), G. Gusman (1998), etc. worked on Araceae found in Pacific and Malaysular region.
As regards classification, H. W. Schott (1860), J. D. Hooker (1883) and J. Hutchinson (1959) divided the family into several tribes where as A. Engler (1879, 1920) divided the family into subfamilies. The later concept has been adopted by most of moden taxonomist such as M. H. Grayum (1991), J. Bogner & D. H. Nicolson (1991), A. Takhtajan (1996), S. J. Mayo & al. (1997), H. Li & al. (2010), etc.
In Vietnam, the botany of Araceae was first studied by J. Loureiro (1790). Later N. E. Brown (1882) described two new species, whereas A. Engler & K. Krause (1911-1920) described many new species from materials collected from Vietnam. However the work of F. Gagnepain (1942) is considered the most systematic and significant with complete record of genera and species. In the last century, Pham Hoang Ho (2000) published his studies on Araceae as the “Illustration in Flora of Southern Vietnam” and “Illustrations in Flora of Vietnam”. Le Kha Ke & al. described common species of Araceae in his book published in 1975. Nguyen Van Du (2005, 2006) compiled the checklist of Araceae in Vietnam. In collaboration with different colleagues he described two genera, many new species and several new records for Vietnam. Besides, several books on the usefulness of the family have also been published from Vietnam by Do Tat Loi (1995), Vo Van Chi (1997, 2012), Do Huy Bich & al. (1993), etc.
Flora of Vietnam – Volume 16 - Araceae is a taxonomic monograph of Araceae in Vietnam. The information presented here are an output of the collections, investigations and research carried out for many years in different regions of Vietnam. To bring out this volume, specimens of the family Araceae were collected in many regions of Vietnam with ecological observations. Specimens stored at several herbaria like Institute of Ecology and Biological Resources (HN), National University of Hanoi (HNU), Institute of Pharmacy (HNIP), Cuc Phuong National Park (CPNP), Institute of Tropical Biology (VNM), Royal Botanic Garden, Kew (K), The Natural History Museum, London (BM), the Phanérogamie, Museum National d'Histoire Naturelle, Paris (P), National Herbarium Nederland, Leiden University Branch (L.), Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universität Berlin (B), University of Copenhagen Herbarium (C), Missouri botanic Garden (MO) and Kunming Institute of Botany (KUN), The Forest herbarium of Department of National Park in Bankok Thailand (BKF), Missouri Botanical Garden, Missouri, USA (MO), etc. were studied. Among these, type specimens have also been checked.
Flora of Vietnam – the family Araceae includes 460 pages. In the book, the author has introduced different systems of classification for Araceae and a system to arrange Araceae taxa in Vietnam. The morphological characters are also analysed carefully. Based on the morphological characters, the keys for genera and species of the family have been prepared. In each taxon, the information on nomenclature, description, type specimens, biology, ecology, distribution in Vietnam and other countries, references, uses and notes (wherever necessary) have been mentioned. Beside description, illustration of each species has been made from the available specimens and cited from other literatures. Photographs are also included 103 line drawings, 97 photos.
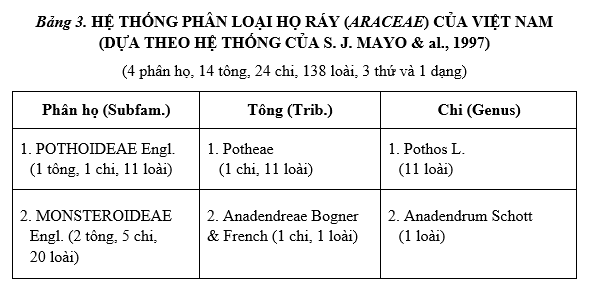
I. CHECKLIST OF THE TAXA IN FAMILY ARACEAE Juss. IN VIETNAM
(4 subfam., 14 trib., 24 gen., 138 sp., 3 var., 1 form.)
Subfam.1. POTHOIDEAE Engl. _ (1 trib., 1 gen., 11 sp.)
Trib.1. POTHEAE (1 gen., 11 sp.)
1. POTHOS L. (11 sp.)
1.1. Pothos pilulifer Buchet ex P. C. Boyce (fig. 10)
1.2. Pothos kerrii Buchet ex P. C. Boyce (fig. 11, photo 1)
1.3. Pothos vietnamensis V. D. Nguyen & P. C. Boyce sp. nov. (photo 3)
1.4. Pothos gigantipes Buchet ex P. C. Boyce (fig. 12, photo 2)
1.5. Pothos chinensis (Raf.) Merr. (fig. 13, photo 4)
1.6. Pothos grandis Buchet ex P. C. Boyce & V. D. Nguyen (fig. 14, photo 5)
1.7. Pothos dzui P. C. Boyce (fig. 15)
1.8. Pothos repens (Lour.) Druce (fig. 16, photo 6)
1.9. Pothos scandens L. (fig. 17, photo 7)
1.10. Pothos lancifolius Hook. f. (fig. 18, photo 8)
1.11. Pothos touranensis Gagnep. (fig. 19)
Subfam.2. MONSTEROIDEAE Engl. (2 trib., 5 gen., 20 sp.)
Trib.2. ANADENDREAE Bogner & French (1 gen., 1 sp.)
2. ANADENDRUM Schott (1 sp.)
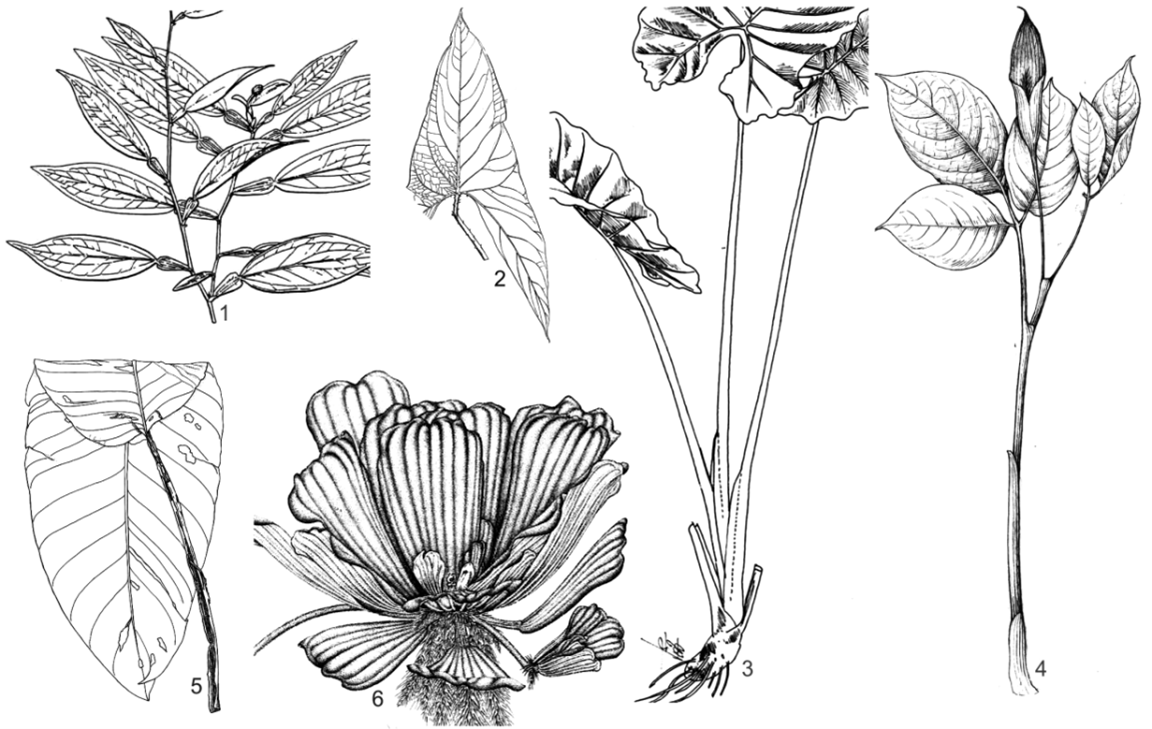
2.1. Anadendrum microstachyum (de Vriese & Miq.) Backer & Alderw. (fig. 20, photo 10)
Trib.3. MONSTEREAE Engl. (4 gen., 19 sp.)
3. RHAPHIDOPHORA Hassk. (13 sp.)
3.1. Rhaphidophora megaphylla H. Li (fig. 21, photo 12)
3.2. Rhaphidophora lacduongensis V. D. Nguyen & B. H. Quang (fig. 22, photo 13)
3.3. Rhaphidophora peepla (Roxb.) Schott (fig. 23)
3.4. Rhaphidophora hookeri Schott (fig. 24, photo 14)
3.5. Rhaphidophora sulcata Gagnep.
3.6. Rhaphidophora bonii Engl. & K. Krause
3.7. Rhaphidophora tonkinensis Engl. & K. Krause (fig. 25)
3.8. Rhaphidophora hongkongensis Schott (fig. 26, photo 15)
3.9. Rhaphidophora chevalieri Gagnep. (fig. 27)
3.10. Rhaphidophora sonlaensis V. D. Nguyen, H. Q. Bui, P. C. Boyce (fig. 28, photo 16)
3.11. Rhaphidophora laichauensis Gagnep. (fig. 29)
3.12. Rhaphidophora decursiva (Roxb.) Schott (fig. 30, photo 17)
3.13. Rhaphidophora crassicaulis Engl. & K. Krause (fig. 31, photo 18)
4. AMYDRIUM Schott (2 sp.)
4.1. Amydrium hainanense (C. C. Ting & C. Y. Wu ex H. Li & al.) H. Li (fig. 32, photo 11)
4.2. Amydrium sinense (Engl.) H. Li (fig. 33, photo 9)
5. EPIPREMNUM Schott (2 sp.)
5.1. Epipremnum giganteum (Roxb.) Schott (fig. 34, photo 19)
5.2. Epipremnum pinnatum (L.) Engl. & K. Krause (fig. 35)
6. SCINDAPSUS Schott (2 sp.)
6.1. Scindapsus officinalis (Roxb.) Schott (fig. 36, photo 20)
6.2. Scindapsus hederaceus (Zoll. & Moritzi) Miq. (fig. 37)
Subfam.3. LASIOIDEAE Engl. (1 trib., 3 gen., 3 sp.)
Trib.4. LASIEAE (3 gen., 3 sp.)
7. PYCNOSPATHA Thorel ex Gagnep. (1 sp.)
7.1. Pycnospatha arietina Gagnep. (photo 21)
8. CYRTOSPERMA Griff. (1 sp.)
8.1. Cyrtosperma merkusii (Hassk.) Schott
9. LASIA Lour. (1 sp.)
9.1. Lasia spinosa (L.) Thw. (fig. 38, photo 22)
Subfam.4. AROIDEAE (10 trib., 15 gen., 104 sp., 3 var., 1 form.)
Trib.5. HOMALOMENEAE (Schott) M. Hotta (1 gen., 5 sp.)
10. HOMALOMENA Schott (5 sp.)
10.1. Homalomena pierreana Engl. & K. Krause (fig. 39)
10.2. Homalomena vietnamense J. Bogner & V. D. Nguyen (photo 23)
10.3. Homalomena occulta (Lour.) Schott (fig. 40, photo 24)
10.4. Homalomena tonkinensis Engl. (fig. 41, photo 25)
10.5. Homalomena pendula (Blume) Bakh. f. (fig. 42)
Trib.6. SCHISMATOGLOTTIDEAE Nakai (1 gen., 2 sp.)
11. SCHISMATOGLOTTIS Zoll. & Moritz. (2 sp.)
11.1. Schismatoglottis harmandii Engl. (fig. 43)
11.2. Schismatoglottis cadieri Buchet & Gagnep. (fig. 44)
Trib.7. AGLAONEMATEAE Engl. (2 gen., 7 sp., 1 form.)
12. AGLAONEMA Schott (6 sp., 1 form.)
Sect.1. CHAMAECAULON Nicolson (2 sp., 1 form.)
12.1. Aglaonema brevispathum (Engl.) Engl. (fig. 45, photo 26)
12.2. Aglaonema costatum N. E. Br.
12.2a. Aglaonema costatum form. immaculatum (Ridl.) Nicolson (photo 27)
Sect.2. AGLAONEMA (4 sp.)
12.3. Aglaonema ovatum Engl.
12.4. Aglaonema modestum Schott ex Engl. (fig. 46, photo 29)
12.5. Aglaonema cochinchinense Engl. (fig. 47)
12.6. Aglaonema simplex Blume (fig. 48, photo 28)
13. AGLAODORUM Schott (1 sp.)
13.1. Aglaodorum griffithii (Schott) Schott (fig. 49)
Trib.8. CALADIEAE Schott (1 gen., 4 sp.)
14. HAPALINE Schott (4 sp.)
14.1. Hapaline locii V. D. Nguyen & Croat (fig. 50, photo 30)
14.2. Hapaline benthamiana Schott (fig. 51)
14.3. Hapaline ellipticifolia C. Y. Wu & H. Li (fig. 52, photo 31)
14.4. Hapaline colaniae Gagnep. (fig. 53)
Trib.9. COLOCASIEAE Engl. (4 gen., 16 sp.)
15. STEUDNERA C. Koch (2 sp.)
15.1. Steudnera colocasiifolia C. Koch (fig. 54, photo 32)
15.2. Steudnera henryana Engl. (fig. 55, photo 33)
16. REMUSATIA Schott (1 sp.)
16.1. Remusatia vivipara (Roxb.) Schott
17. COLOCASIA Schott (5 sp.)
17.1. Colocasia menglaensis J. T. Yin, H. Li & Z. F. Xu (fig. 56, photo 34)
17.2. Colocasia esculenta (L.) Schott (photo 35)
17.3. Colocasia gigantea (Blume) Hook. f. (photo 37)
17.4. Colocasia lihengiae C. L. Long & K. M. Liu (photo 38)
17.5. Colocasia yunnanensis C. L. Long & X. Z. Cai. (photo 39)
18. ALOCASIA (Schott) G. Don (8 sp.)
18.1. Alocasia macrorrhizos (L.) G. Don (photo 40)
18.2. Alocasia cucullata (Lour.) G. Don (photo 36)
18.3. Alocasia longiloba Miq. (fig. 57, photo 41)
18.4. Alocasia odora (Roxb.) C. Koch (fig. 58, photo 42)
18.5. Alocasia decumbens Buchet
18.6. Alocasia evrardii Gagnep. ex V. D. Nguyen (fig. 59, photo 43)
18.7. Alocasia vietnamensis V. D. Nguyen & de Kok (fig. 60, photo 44)
18.8. Alocasia lecomtei Engl.
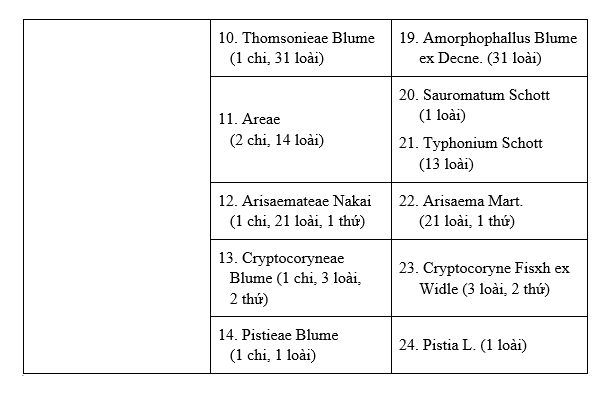
Trib.10. THOMSONIEAE Blume (1 gen., 31 sp.)
19. AMORPHOPHALLUS Blume ex Decne. (31 sp.)
19.1. Amorphophallus opertus Hett. (fig. 61, photo 45)
19.2. Amorphophallus scaber Serebryanyi & Hett. (photo 46)
19.3. Amorphophallus paeoniifolius (Dennst.) Nicolson (fig. 62, photo 47)
19.4. Amorphophallus pusillus Hett. & Serebryanyi (fig. 63)
19.5. Amorphophallus coaetaneus S. Y. Liu & S. J. Wei (fig. 64, photo 48)
19.6. Amorphophallus hayi Hett. (fig. 65)
19.7. Amorphophallus rhizomatosus Hett. (fig. 66, photo 49)
19.8. Amorphophallus pilosus Hett. (fig. 67, photo 50)
19.9. Amorphophallus lanuginosus Hett. (fig. 68, photo 51)
19.10. Amorphophallus konjac K. Koch (photo 52)
19.11. Amorphophallus interruptus Engl. & Gehrm. (fig. 69, photo 53)
19.12. Amorphophallus orchroleucus Hett. & V. D. Nguyen (fig. 70, photo 54)
19.13. Amorphophallus kienluongensis V. D. Nguyen, Luu & Hett. (photo 55)
19.14. Amorphophallus dzui Hett. (fig. 71, photo 56)
19.15 Amorphophallus sinuatus Hett. & V. D. Nguyen (fig. 72, photo 57)
19.16. Amorphophallus glossophyllus Hett. (photo 58)
19.17. Amorphophallus tenuistylis Hett. (photo 60)
19.18. Amorphophallus harmandii Engl. & Gehrm (fig. 73)
19.19. Amorphophallus coudercii (Bogner) Bogner (fig. 74, photo 59)
19.20. Amorphophallus corrugatus N. E. Br. (fig. 75, photo 61)
19.21. Amorphophallus krausei Engl. & Gehrm.
19.22. Amorphophallus synandrifer Hett. & V. D. Nguyen (fig. 76)
19.23. Amorphophallus verticillatus Hett. (fig. 77, photo 62)
19.24. Amorphophallus mekongensis Engl. & Gehrm.
19.25. Amorphophallus yunnanensis Engl. & Gehrm. (fig. 78, photo 63)
19.26. Amorphophallus tonkinensis Engl. & Gehrm. (fig. 79, photo 64)
19.27. Amorphophallus fallax (Serebryanyi) Hett. & Claudel
19.28. Amorphophallus pseudoharmandii Hett. & Claudel (fig. 80)
19.29. Amorphophallus lacourii Linden & André (fig. 81, photo 65)
19.30. Amorphophallus kuznetsovii (Serebryanyi) Hett. & Claudel (photo 66)
19.31. Amorphophallus lanceolatum (Serebryanyi) Hett. & Claudel
Trib.11. AREAE (2 gen., 14 sp.)
20. SAUROMATUM Schott (1 sp.)
20.1. Sauromatum horsfieldii Miq. (fig. 82)
21. TYPHONIUM Schott (13 sp.)
21.1. Typhonium flagelliforme (Lodd.) Blume (fig. 83, photo 67)
21.2. Typhonium lineae Hett. & V. D. Nguyen (fig. 84, photo 68)
21.3. Typhonium stigmatilobatum V. D. Nguyen (fig. 85, photo 69)
21.4. Typhonium huense V. D. Nguyen & T. Croat (photo 70)
21.5. Typhonium acetosella Gagnep. (fig. 86)
21.6. Typhonium bachmaense V. D. Nguyen & Hett. (fig. 87, photo 71)
21.7. Typhonium violifolium Gagnep.
21.8. Typhonium vermiforme V. D. Nguyen & T. Croat (photo 72)
21.9. Typhonium circinnatum Hett. & J. Mood (photo 73)
21.10. Typhonium penicillatum V. D. Nguyen & Hett. (fig. 88, photo 74)
21.11. Typhonium blumei Nicolson & Sivadasan (photo 75)
21.12. Typhonium hayatae Sriboonma & Murata
21.13. Typhonium trilobatum (L.) Schott (photo 76)
Trib.12. ARISAEMATEAE Nakai (1 gen., 21 sp., 1 var.)
22. ARISAEMA Mart. (21 sp., 1 var.)
Sect.1. Anomala G. & L. Gusman (6 sp.)
22.1. Arisaema balansae Engl. (fig. 89, photo 77)
22.2. Arisaema victoriae V. D. Nguyen (fig. 90, photo 78)
22.3. Arisaema pingbianense H. Li (fig. 91, photo 80)
22.4. Arisaema rostratum Nguyen V. D. & P. C. Boyce (photo 81)
22.5. Arisaema omkoiense Gusman (fig. 92)
22.6. Arisaema petelotii K. Krause (fig. 93, photo 82)
Sect.2. Fimbriata Engl. (6 sp.)
22.7. Arisaema garrettii Gagnep. (fig. 94, photo 83)
22.8. Arisaema petiolulatum Hook. F. (fig. 95)
22.9. Arisaema claviforme P. Bruggeman & al. (photo 79)
22.10. Arisaema averyanovii V. D. Nguyen & P. C. Boyce (fig. 96, photo 84)
22.11. Arisaema pierreanum Engl. (photo 85)
22.12. Arisaema condaoense V. D. Nguyen
Sect.3. Tortuosum (4 sp.)
22.13. Arisaema ramulosum Alderw. (fig. 97, photo 86)
22.14. Arisaema lidaense J. Murata & S. G. Wu
22.15. Arisaema honbaense Luu, Tich, G. Tran & V. D. Nguyen (photo 87)
22.16. Arisaema roxburghii Kunth (fig. 98, photo 88)
Sect.4. Decipientia Engl. (2 sp., 1 var.)
22.17. Arisaema decipiens Schott (fig. 99, photo 89)
22.18. Arisaema rhizomatum C. E. C. Fisch (fig. 100: 1, 4, 5; photo 90: 1).
22.18a. Arisaema rhizomatum var. nudum C. E. C. Fisch. (fig. 100: 2, 3; photo 90: 2, 3, 4)
Sect.5. Franchetiana Engl. (1 sp.)
22.19. Arisaema franchetianum Engl. (fig. 101, photo 91)
Sect.6. Sinarisaema Nakai (2 sp.)
22.20. Arisaema consanguineum Schott (fig. 102, photo 92)
22.21. Arisaema parisifolia J. Murata
Trib.13. CRYPTOCORYNEAE Blume (1 gen., 3 sp., 2 var.)
23. CRYPTOCORYNE Fisch. ex Widle (3 sp., 2 var.)
23.1. Cryptocoryne crispatula Engl.
23.1a. Cryptocoryne crispatula var. balansae (Gagnep.) N. Jacobsen (photo 93)
23.1b. Cryptocoryne crispatula var. tonkinensis (Gagnep.) N. Jacobsen (photo 94)
23.2. Cryptocoryne annamica Serebryanyi (photo 95)
23.3. Cryptocoryne vietnamensis Hertel & Muhlberg (fig. 103, photo 96)
Trib.14. PISTIEAE Blume (1 gen., 1 sp.)
24. PISTIA L. (1 sp.)
24.1. Pistia stratiotes L. (photo 97)
III. KEY TO THE GENERA OF FAMILY ARACEAE IN VIETNAM
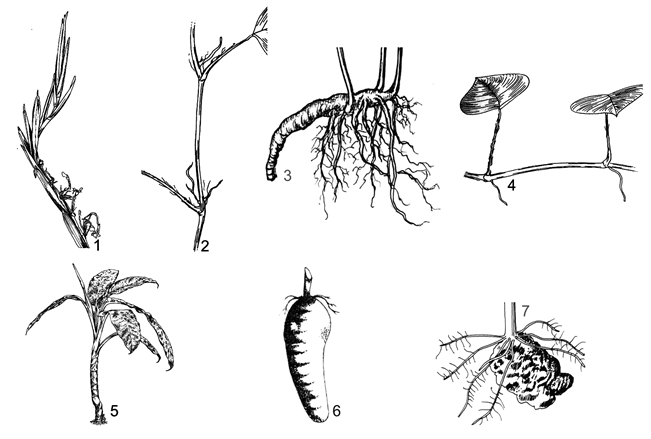
| 1A. |
Plants grow on ground, epiphyte on trees, rocks, rarely equatic (aquatic but not floating); with or without perianth, spadix bearing many bisexual or monosexual flowers. |
|
| 2A. |
Spadix bearing bisexual flowers, not divided into different portions, flowers with or without perianth. |
|
| 3A. |
Epiphyte lianas creeping on ground or climbing on trees; without spines on stems and leaves. |
|
| 4A. |
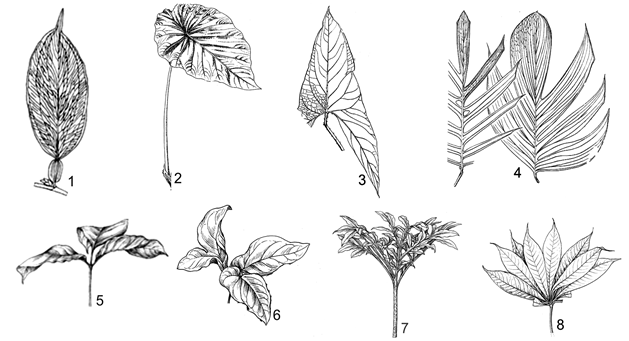
Flowers with free perianth, 4-6 pieces; ovary 3 cells; 1 ovule in each; leaf blade entire, lateral vein archly forward to apex from leaf blade base; petiole flattened as wings |
(Subfam.1. POTHOIDEAE, Trib.1. Potheae)
1. POTHOS
|
| 4B. |
Flowers naked or with compound perianth as cup form; ovary 1 rarely 2 cells; ovules 2-∞, rarely 1; leaf blade entire to pinnatisect, veins pinnative; petioles more or less sheathed. |
(Subfam.2. MONSTEROIDEAE). |
| 5A. |
With compound perianth like cup form; ovary 1 cell, 1 ovule; vegetative organs without clerochleid; sheaths soon dried but not remain fibres. |
(Trib.2. Anadendreae) 2. ANADENDRUM |
| 5B. |
Without perianth; ovaries 1, rarely 2 cells; ovules 2-∞, seldom 1 ovule; vegerative organs with abundant clerochleid; leaf blades entire, divided, pinnatisect or perforated; sheaths obvious or non obvious, soon dried and remain fibres. |
(Trib.3. Monstereae). |
| 6A. |
Ovules abundant, with long stipes. |
3. RHAPHIDOPHORA |
| 6B. |
Ovules poor (1-6), with short stipe. |
|
| 7A. |
Ovules 2-4(6); petiole sheath soon deciduous or dried. |
|
| 8A. |
Petiole sheath soon deciduous, veination prominent from leaf beneath surface |
4. AMYDRIUM |
| 8B. |
Petiole sheath dried, remain fibres, veinations not prominent from leaf beneath surface |
5. EPIPREMNUM |
| 7B. |
Ovule 1; petiole sheaths existent |
6. SCINDAPSUS |
| 3B. |
Rhizomatous or tuberous plants, bearing spines on organs; growing on ground or in swampes or marshes |
(Subfam.3. LASIOIDEAE; Trib.4. Lasieae). |
| 9A. |
Growing in dry places; leaf blades dracontioid shape; inflorescences short, total 15-20 cm long. |
7. PYCNOSPATHA |
| 9B. |
Growing in swampers or marshes; leaf blades entire or pinnatisect; inflorescence long, total up to longer 20 cm. |
|
| 10A. |
Leaf blades having posterior lobes longer than anterior lobes, ovule abundant, pariental; tuberous plant. |
8. CYRTOSPERMA |
| 10B. |
Leaf blade having posterior lobes shorter than anterior lobes; ovule 1, apical; rhizomatous plants. |
9. LASIA |
| 2B. |
Spadix divided into male, female or sterile portions |
(Subfam.4. AROIDEAE). |
| 11A. |
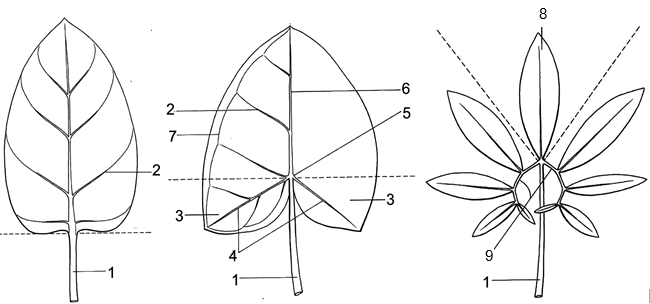
On leaf blades, small veins parallely forward to margin from midrib; lateral veins not connected together by a collective vein. |
|
| 12A. |
Rhizomes more or less romatic; spathes not seperated into tube and lamina, common in green color, only open in anthesis periods, existent with female flowers; ovaries 2-4 cells; ovule abundant. |
(Trib.5. Homalomeneae)
10. HOMALOMENA
|
| 12B. |
Rhizomes without romatic; spathes seperated or not seperated into tube and lamina, total spathe or only lamina soon deciduous after synthesis, tube existent with female flowers; ovaries 1-2 cells; 1 ovule in each. |
|
| 13A. |
Spathes seperated into tube and lamina, only laminas opened in synthesis period then soon deciduous, tubes not open, existent. |
(Trib.6. Schismatoglottideae)
11. SCHISMATOGLOTTIS
|
| 13B. |
Spathes not seperated into tube and lamina, total spathe opened in synthesis period then deciduous. |
(Trib.7. Aglaonemateae). |
| 14A. |
Growing on ground; stems earial, straight, sometime creeping; pduncle often shorter than petiole; spadix with several female flower rings at base. |
12. AGLAONEMA |
| 14B. |
Aquatic plant; stem creeping rarely straightward; peduncles subequal to petioles; spadix with 1 female flower ring at base. |
13. AGLAODORUM |
| 11B. |
On leaf blade, small veins forward from lateral veins, reticulately connect together; lateral veins connected together by collective vein. |
|
| 15A. |
Growing on ground, wet places but never as equatic plants; spadix appendix free from spathe. |
|
| 16A. |
Filaments connected into columns or board; anthers hidden. |
|
| 17A. |
Leaf blades varied but never peltate; ovaries 1 cell; ovule 1. |
(Trib.8. Calladieae) 14. HAPALINE |
| 17B. |
Leaf blades more or less seltate; ovaries 1-4 cells; ovules several to abundant. |
(Trib.9. Colocasieae) |
| 18A. |
Spadix without appendix at apex. |
|
| 19A. |
Between ovaries having staminodes, without propagating stolons. |
15. STEUDNERA |
| 19B. |
Between ovaries not having staminodes; with propagating stolons. |
16. REMUSATIA |
| 18B. |
Spadix often rarely without with appendix portion at apex. |
|
| 20A. |
Ovules abundant, ariental placentas; small veins conspicuous. |
17. COLOCASIA |
| 20B. |
Ovules several, basal placentas; small veins nonconspicuous. |
18. ALOCASIA |
| 16B. |
Filament not connected into column or boards; anthers conspicuous. |
|
| 21A. |
Petiole sheaths often very short or not conspicuous, leaf blades in dracontioid shape; spadix often with stout appendix, rarely slender or subulate. |
(Trib.10. Thomsonieae). 19. AMORPHOPHALLUS |
| 21B. |
Petiole sheaths conspicuous; leaf blade entire, trifoliate, radiate; appendix often slender, rarely stout. |
|
| 22A. |
Spadix bisexual, having sterile portion between male and female portions |
(Trib.11. Areae). |
| 23A. |
Leaf blade palmate, not constricted into tube and lamina, sterile portion between male and female portion not having nake axile portion. |
20. SAUROMATUM |
| 23B. |
Leaf blade entire or trilobed, rarely palmate lobed; spathe constricted at middle and separate into tube and lamina, sterile portion between male and female portion having nake axile portion (not including T. flagelliforme). |
21. TYPHONIUM |
| 22B. |
Spadix mono sexual and bisexul, if bisexul not having sterile portion between male and female portion. |
(Trib.12. Arisaemateae)
22. ARISAEMA
|
| 15B. |
Aquatic plant in rivers or streamlines; spadix appendix attached with spathe. |
(Trib.13. Cryptocoryneae)
23. CRYPTOCORYNE
|
| 1B. |
Floating aquatic plant; leaves spathulate, petioles not conspicuous, rosette in arrange; spadix bearing only 1 ovary, 2-8 male flowers. |
(Trib.14. Pistieae)
24. PISTIA
|
III. KEY TO THE SPECIES OF FAMILY ARACEAE IN VIETNAM
3.1. KEY TO THE SPECIES OF THE GENUS POTHOS IN VIETNAM
| 1A. |
Petiole flattened as a broad wing |
|
| 2A. |
Peduncle more or less straight upward. |
|
| 3A. |
Spadix globose or ovate. |
|
| 4A. |
Spadix small, < 5 mm diameter. |
|
| 5A. |
Peduncle shorter (< 5 mm), full covered by cataphylls. |
1. P. pilulifer |
| 5B. |
Peduncle longer (> 2 cm), only base covered by cataphylls. |
|
| 6A. |
Spadix stipitated, globose, c. 5mm diam. |
2. P. kerrii |
| 6B. |
Spadix sessile, ovate or slightly cylindrical, 3-3.5 mm diam. |
3. P. vietnamensis |
| 4B. |
Spadix quite large, > 7 mm diam. |
|
| 7A. |
Spadix globose. |
4. P. gigantipes |
| 7B. |
Spadix elliptic or obovate. |
|
| 8A. |
Peduncle short; spadix elliptic or slight ovate. |
5. P. chinensis |
| 8B. |
Peduncle long; spadix obovate. |
6. P. grandis |
| 3B. |
Spadix cylindrical, long. |
|
| 9A. |
Spadix stout cylindrical; petioles much shorter than leaf laminas. |
7. P. dzui |
| 9B. |
Spadix slender cylindrical; petioles longer than leaf laminas. |
8. P. repens |
| 2B. |
Peduncle bend down on stem. |
9. P. scandens |
| 1B. |
Petiole not as obove. |
|
| 10A. |
Peduncle 1.5-5 mm long; spathe lanceolate, size 2.5-10 × 1-4 cm; spadix sessile. |
10. P. lancifolius |
| 10B. |
Peduncle 10-19 mm long; spathe triangular, size 6-9 × 1,5-2 cm; spadix with stipes |
11. P. touranensis |
3.2. KEY TO THE SPECIES OF GENUS ANADENDRUM IN VIETNAM
| With compound perianth like cup form; ovary 1 cell, 1 ovule; vegetative organs without clerochleid; sheaths soon dried but not remain fibres. |
1. A. microstachyum |
3.3. KEY TO THE SPECIES OF GENUS RHAPHIDOPHORA IN VIETNAM
| 1A. |
Leaf blades entire. |
|
| 2A. |
Leaf blade large, ovate to broad ovate. |
|
| 3A. |
Leaf blade base cordate, width subequal length; spadix large, 12-14 cm long; ovary with long style. |
1. R. megaphylla |
| 3B. |
Leaf blade base round, length > width obviously; spadix max. 8 cm long; ovary concaved at surface, style not obvisiously seen. |
2. R. lacduongensis |
| 2B. |
Leaf blade almost small, lanceolate to elongate elliptic or ovate, width much shorter than length, base acute, obtus or round. |
|
| 4A. |
Petiole equal or longer than leaf blade, leaf base always round. |
|
| 5A. |
Stigma elliptic, usually constricted into 8 shape, parallel with spadix axile; leaf blade ovate to falcate ovate, lateral vein not much prominent from beneath surface. |
3. R. peepla |
| 5B. |
Stigma round; leaf blade always elliptic, lateral veins much prominent from beneath surface |
4. R. hookeri |
| 4B. |
Petiole much shorter than leaf blade; leaf base obtuse or acute. |
|
| 6A. |
Leaf blade small, length always < 15 cm; stem angulated, narrow winged. |
5. R. sulcata |
| 6B. |
Leaf blade not small, length > 15 cm; stem cylindrical, not winged. |
|
| 7A. |
Lateral veins upward by large angle (60-75o) from midrib. |
|
| 8A. |
Style region larger than ovary, surface hexagonal; stamens opened by basal slits |
6. R. bonii |
| 8B. |
Style region smaller or equal ovary, surface tetragonal; stamens opened by slit at surface |
7. R. tonkinensis |
| 7B. |
Lateral veins upward by narrow angle (< 50o) from midrib. |
|
| 9A. |
Placentas axile, flattened, sub equal ovary in width; ovule abundant, attached on placenta surface; stamens opened by long slit on surface. |
8. R. hongkongensis |
| 9B. |
Placentas axile, slender; ovules attached at base placenta; stamens opened by short slits at base. |
9. R. chevalieri |
| 1B. |
Adult leaf blade pinnately divided, entire blade only in juvenile period. |
|
| 10A. |
Leaf blade small, < 40 cm long, leaf lobes several (< 5); spadix small, < 12 cm long. |
|
| 11A. |
Petiole sheath broken into fragment flat tissues when decayed; spadix slender cylindrical; stigma elliptic, not concave. |
10. R. sonlaensis |
| 11B. |
Petiole sheath broken into fibres when decayed; spadix conical; stigma round, concave |
11. R. laichauensis |
| 10B. |
Leaf blade large, > 40 cm long, leaf lobes many (> 7); spadix large, > 12 cm long. |
|
| 12A. |
Leaf lobes without perforation; spadix cylindrical, stout, straight, surface even; stem cylindrical |
12. R. decursiva |
| 12B. |
Leaf lobes with perforation; spadix light conical, always slight bend down, surface uneven |
13. R. crassicaulis |
3.4. KEY TO THE SPECIES OF GENUS AMYDRIUM IN VIETNAM
| 1A. |
Leaf blade with perforation, base cordate. |
1. A. hainanense |
| 1B. |
Leaf blade without perforation, base obtuse to acute. |
2. A. sinense |
3.5. KEY TO THE SPECIES OF GENUS EPIPREMNUM IN VIETNAM
| 1A. |
Leaf blades entire. |
1. E. giganteum |
| 1B. |
Leaf blades devided pinnately. |
2. E. pinnatum |
3.6. KEY TO THE SPECIES OF GENUS SCINDAPSUS IN VIETNAM
| 1A. |
Stigma elliptic, prominent from surface; leaves broad ovate. |
1. S. officinalis |
| 1B. |
Stigma linear, concaved at middle; leaves ovate to lanceolate. |
2. S. hederaceus |
3.7. KEY TO THE SPECIES OF GENUS PYCNOSPATHA IN VIETNAM
| Growing in dry places; leaf blades dracontioid shape; inflorescences short, total 10-20 cm long. |
1. P. arietina |
3.8. KEY TO THE SPECIES OF GENUS CYRTOSPERMA IN VIETNAM
| Leaf blades having posterior lobes longer than anterior lobes, ovule abundant, pariental; tuberous plant |
1. C. merkusii |
3.9. KEY TO THE SPECIES OF GENUS LASIA IN VIETNAM
| Leaf blade having posterior lobes shorter than anterior lobes; ovule 1, apical; rhizomatous plants |
1. L. spinosa |
3.10. KEY TO THE SPECIES OF GENUS HOMALONEMA IN VIETNAM
| 1A. |
Base of leaf blade not concave, without posterior lobes. |
|
| 2A. |
Leaf blade lanceolate to elongate triangular, width < 4 cm, base truncate to obtuse or acute. |
1. H. pierreana |
| 2B. |
Leaf blade ovate to elliptic, width > 4 cm, base round. |
2. H. vietnamense |
| 1B. |
Base of leaf blade concave less or more cordate, posterior lobes conspicuous. |
|
| 3A. |
Small to medium rhizomatous herb, 30-50 cm tall; leaf blade seldom < 35 cm; spathe length < 6.5 cm. |
|
| 4A. |
Pariental placenta; without style; staminodes in female portion globose at apex, stipe long |
3. H. occulta |
| 4B. |
Axile placenta; style conspicuous; staminodes in female portion truncate clavate, stipe short |
4. H. tonkinensis |
| 3B. |
Rather big rhizomatous herb, to 1 m tall; leaf blade to 50 cm long; spathe length > 10 cm. |
5. H. pendula |
3.11. KEY TO THE SPECIES OF GENUS SCHISMATOGLOTTIS IN VIETNAM
| 1A. |
Spathe tube longer than 1/2 spathe lamina; male portion capitate or subclavate |
1. S. harmandii |
| 1B. |
Spathe tube shorter than 1/2 spathe lamina; male portion cylindrical to subconical. |
2. S. cadieri |
3.12. KEY TO THE SPECIES OF GENUS AGLAONEMA IN VIETNAM
| 1A. |
Stem branched, creeping on ground; petiole sheath shorter 1cm. (Sect.1. Chamaecaulon) |
|
| 2A. |
With dried cataphylls remaining; petiole equal or longer than leaf blade; leaf base obtus or round |
1. A. brevispathum |
| 2B. |
Without dried cataphyll; petiole shorter than leaf blade; leaf base round to cordate. |
2. A. costatum
2a. A. costatum form. immaculatum
|
| 1B. |
Stem rarely branched, upward not creeping on ground; petiole sheath longer 1 cm. (Sect.2. Aglaonema) |
|
| 3A. |
Spadix stipe and female portion attached to spathe. |
|
| 4A. |
Between female and male portion with sterile portion; peduncle 3.5-7 cm long. |
3. A. ovatum |
| 4B. |
Female portion continued be male portion, without sterile portion; peduncle 5-10 cm long. |
4. A. modestum |
| 3B. |
Spadix stipe and female portion free from spathe.Spadix stipe and female portion free from spathe. |
|
| 5A. |
Lateral veins not conspicuous.Lateral veins not conspicuous. |
5. A. cochinchinense |
| 5B. |
Lateral veins conspicuous.Lateral veins conspicuous. |
6. A. simplex |
3.13. KEY TO THE SPECIES OF GENUS AGLAODORUM IN VIETNAM
| Equatic plant; stem creeping rarely straightward; peduncles subequal to petioles; spadix with 1 female flower ring at base. |
1. A. griffithii |
3.14. KEY TO THE SPECIES OF GENUS HAPALINE IN VIETNAM
| 1A. |
Spadix appendix confused to sterile half of male flowers; leaf blade bases cordate, posterior lobes not really distinguished from blade. |
1. H. locii |
| 1B. |
Spadix appendix seperated from male flowers; leaf blade base deeply concaved, posterior lobes distinguished from blade. |
|
| 2A. |
Leaf blade ovate to elliptic, not constricted. |
|
| 3A. |
Leaf blade ovate to broad ovate, posterior lobes outward; ovaries ovate. |
2. H. benthamiana |
| 3B. |
Leaf blade elliptic, posterior lobe nearly parallel together; ovaries bottle shape. |
3. H. ellipticifolia |
| 2B. |
Leaf blade sagitate, anterior lobe constricted at base. |
4. H. colaniae |
3.15. KEY TO THE SPECIES OF GENUS STEUDNERA IN VIETNAM
| 1A. |
Spathe lamina light brown outside, purple inside; pariental placenta. |
1. S. colocasifolia |
| 1B. |
Spathe lamina yellow outside, red-brown inside; base placenta... |
2. S. henryana |
3.16. KEY TO THE SPECIES OF GENUS REMUSATIA IN VIETNAM
| Between ovaries not having staminodes; with propagating stolons. |
1. R. vivipara |
3.17. KEY TO THE SPECIES OF GENUS COLOCASIA IN VIETNAM
| 1A. |
Spadix with appendix longer than 3 cm. |
|
| 2A. |
Petiole and peduncle rubescence; spathe without fragrant, dark yellow. |
1. C. menglaensis |
| 2B. |
Petiole and peduncle glabrous; spathe fragrant, light yellow. |
2. C. esculenta |
| 1B. |
Spadix with or without appendix, if with, appendix small and shorter 2 cm. |
|
| 3A. |
Spathe blade white; appendix small, shorter 2 cm long; petiole waxy-white glaucous |
3.C. gigantea |
| 3B. |
Spathe blade yellow; appendix absent; petiole glabrous. |
|
| 4A. |
Leaf blade without dark purple area, basal lobes triangular shape. |
4. C. lihengiae |
| 4B. |
Leaf blade with dark purple area, basan lobes round. |
5. C. yunnanensis |
3.18. KEY TO THE SPECIES OF GENUS ALOCASIA IN VIETNAM
| 1A. |
Leaf blade without any connection between posterior lobes; inflorescence big > 20 cm; spathe lamina reflect spadix axil; appendix long >5 cm, straight. |
1. A. macrorrhizos |
| 1B. |
Leaf blade less or more with connection between posterior lobes; inflorescence smaller above; spathe lamina cymbiform, straight or arch; appedndix smaller than above, a little curved. |
|
| 2A. |
Lateral veins almost forward from leaf blade base, posterior lobes round. |
2. A. cucullata |
| 2B. |
Lateral vein forward from midrib, posterior lobes acute, obtuse or round. |
|
| 3A. |
Posterior lobes of leaf blade with connection < 1/2 their length. |
|
| 4A. |
Posterior lobes triangular, acute at apex, equal 1/2 anterior lobe; lateral veins 3-4 pairs |
3. A. longiloba |
| 4B. |
Posterior lobes semi-elliptic, obtus or round at apex, < 1/2 anterior lobe; leteral veins 4-10 pairs. |
|
| 5A. |
Spadix with female portion conical, >2,5 cm long, male portion much longer than sterile portion. |
4. A. odora |
| 5B. |
Spadix with female portion conical, < 2 cm long, male portion slight conical, shorter than sterile portion. |
5. A. decumbens |
| 3B. |
Posterior lobes with connection > 1/2 their length. |
|
| 6A. |
Appendix stipitate. |
|
| 7A. |
Tuberous plant. |
6. A. evrardii |
| 7B. |
Rhizomatous plant. |
7. A. vietnamensis |
| 6B. |
Appendix sessile. |
8. A. lecomtei |
3.19. KEY TO THE SPECIES OF GENUS AMORPHOPHALLUS IN VIETNAM
| 1A. |
Stamens free, filament short, no obviously; appendix surface not brain surface like |
|
| 2A. |
Peduncle shorter or equal spathe. |
|
| 3A. |
PSpadix shorter than spathe, hiden in spathe at synthesis period. |
|
| 4A. |
Ovaries 1 cell; stigma non divided to lobes. |
1. A. opertus |
| 4B. |
Ovaries 2 cells; stigma divided to lobes. |
2. A. scaber |
| 3B. |
Spadix equal spathe or longer, extrosed from spathe in synthesis period. |
|
| 5A. |
Inflorescence big to huge; appendix conical, concaved and varied shapes. |
3. A. paeoniifolius |
| 5B. |
Inflorescence small; appendix subulate. |
4. A. pusillus |
| 2B. |
Peduncle longer than spathe. |
|
| 6A. |
Spadix much longer than spathe. |
|
| 7A. |
Leaves and inflorescence present together. |
|
| 8A. |
Evergreen tuberous plants; appendix elongate conical, straight. |
5. A. coaetaneus |
| 8B. |
Deciduous rhizomatous plants; appendix long subulate, ben down. |
|
| 9A. |
Spadix stipitate; appendix wrinkled and bearing staminodes. |
6. A. hayi |
| 9B. |
Spadix sessile; appendix not as above. |
7. A. rhizomatosus |
| 7B. |
Leaves and inflorescence present in different times. |
|
| 10A. |
Spadix bearing hairy sterile projections. |
|
| 11A. |
Black hairy projections present on appendix. |
8. A. pilosus |
| 11B. |
Pale green or lighy yellow hairy projection on fertile portions. |
9. A. lanuginosus |
| 10B. |
Spadix without hairy sterile projection. |
|
| 12A. |
Tuber less or more pressed globose. |
|
| 13A. |
Inflorescence large, longer than 1 m, appendix dark meat red. |
10. A. konjac |
| 13B. |
Inflorescence much smaller, never upto 1 m, appendix white or cream color. |
|
| 14A. |
Stamens arranged in horizonal bands on spadix axil. |
11. A. interruptus |
| 14B. |
Stamens arranged dense but not in bands in horizonal. |
|
| 15A. |
Spathe light brown with darker brown spots; appendix oblong, obtuse at apex. |
12. A. orchroleucus |
| 15B. |
Spathe light green to white; appendix conical, acute at apex. |
13. A. kienluongensis |
| 12B. |
Tubers elongate. |
|
| 16A. |
Spadix with staminode portion between male and female portions. |
14. A. dzui |
| 16B. |
Spadix without staminode portion between male and female portions. |
|
| 17A. |
Peduncle shorter than petiole; spathe short only covers female portion. |
15. A. sinuatus |
| 17B. |
Peduncle equal, rarely shorter petiole; spathe longer covers both female and male portion. |
|
| 18A. |
Stigma discoid. |
16. A. glossophyllus |
| 18B. |
Stigma 2 or 3 lobes. |
|
| 19A. |
Plant up to 1 m high; inflorescence longer than 1 m. |
17. A. tenuistylis |
| 19B. |
Plant much smaller, lower 50 cm; inflorescence shorter than 40 cm. |
18. A. harmandii |
| 6B. |
Spadix shorter, equal or slightly extrude from spathe. |
|
| 20A. |
Spadix without appendix. |
19. A. coudercii |
| 20B. |
Spadix always with appendix. |
|
| 21A. |
Appendix wrinkled as brain surface. |
20. A. corrugatus |
| 21B. |
Appendix not resemble brain surface. |
|
| 22A. |
Spadix bearing staminode portion between male and female portions. |
|
| 23A. |
Stigma round, smaller than ovaries. |
21. A. krausei |
| 23B. |
Stigma elliptic to lozenge, larger than ovaries. |
22. A. synandrifer |
| 22B. |
Spadix without staminode. |
|
| 24A. |
Rhizomatous stem, much branched; growing in cluster. |
23. A. verticillatus |
| 24B. |
Tuberous stem, unbranched; growing solitary. |
|
| 25A. |
Without style. |
24. A. mekongensis |
| 25B. |
With styles. |
|
| 26A. |
Spadix with stipe; appendix surface concaved. |
25. A. yunnanensis |
| 26B. |
Spadix without stipe. Appdendix surface not concaved. |
26. A. tonkinensis |
| 1B. |
Stamens free, filament long, obviously; appendix surface resembles brain surface. |
|
| 27A. |
Style very short or non conspicuous. |
|
| 28A. |
Appendix with conspicuous stipe (> 5 mm), surface resemble brain; peduncle longer than petiole. |
27. A. fallax |
| 28B. |
Appendix sessile (< 5 mm), surface covered by clavate sterile projections; peduncle shorter than petiole. |
28. A. pseudoharmandii |
| 27B. |
Style long, conspicuous. |
|
| 29A. |
Plants grow in bluster, leaves spotted; peduncle shorter or equal petiole; male flowers arrange dense. |
29. A. lacourii |
| 29B. |
Plants grow solitary, leaves not spotted; peduncle longer than petiole; male flowers arrange scattered. |
|
| 30A. |
Plant > 70 cm tall; petiole brown; appendix and male portion flattened at dossal; spadix > 20 cm long. |
30. A. kuznetsovii |
| 30B. |
Plant < 60 cm; petiole green; appendix and male portion not flattened; spadix < 8 cm |
31. A. lanceolatum |
3.20. KEY TO THE SPECIES OF GENUS SAUROMATUM IN VIETNAM
| Leaf blade palmate, not constricted into tube and lamina, sterile portion between male and female portion not having nake axile portion. |
1. S. horsfieldii |
3.21. KEY TO THE SPECIES OF GENUS TYPHONIUM IN VIETNAM
| 1A. |
On the spadix, sterile portion between male and female portions with 2 different sterile flower styles, clavate form at lower and subulate at at upper. |
1. T. flagelliforme |
| 1B. |
Spadix bearing only 1 sterile flower style. |
|
| 2A. |
Spathe lamina elongate narrow laceolate or triangular, thin. |
|
| 3A. |
Sterile flower short, less or more cylindrical, clavate or broad linear. |
|
| 4A. |
Leaf blade pedate, lobes narrow lanceolate or linear, maximum < 1.3 cm. |
2. T. lineae |
| 4B. |
Leaf blade entire, trifoliate or more, lobes ovate to triangular or broad lanceolate, minimum > 1.5 cm. |
|
| 5A. |
Stigma concave, shallowly lobed. |
3. T. stigmatilobatum |
| 5B. |
Stigma discoid, not concaved and lobed. |
|
| 6A. |
Sterile flowers lozenge, acute at both apexes. |
4. T. huense |
| 6B. |
Sterile flowers clavate. |
5. T. acetosella |
| 3B. |
Sterile flowers filiforms or slender subulate, 3-6 mm long, curved. |
|
| 7A. |
Sterile flowers clavate at apex, lower part filiform |
6. T. bachmaense |
| 7B. |
Sterile flowers filiform but not clavate at apex. |
|
| 8A. |
Spathe 13 cm long, purple, sterile flowers straight. |
7. T. violaliforme |
| 8B. |
Spathe longer than 15 cm, not red, sterile flowers curved. |
8. T. vermiforme |
| 2B. |
Spathe lamina triangular to broad ovate, rather thickened. |
|
| 9A. |
Sterile flowers broad clavate; spathe convoluted. |
9. T. circinnatum |
| 9B. |
Sterile flowers filiform; spathe not as above. |
|
| 10A. |
Sterile flowrs less or more straight, not curved together. |
|
| 11A. |
Plant with entire and trifoliate leaves together; stigma penicillate on surface. |
10. T. penicillatum |
| 11B. |
Plant with entire leaves only, stigma not penicillate on surface. |
|
| 12A. |
Sterile flowers filiform, to 7 mm long, straight, dense. |
11. T. blumei |
| 12B. |
Sterile flowers less or more cylindric, 1-2 mm long, scattered. |
12. T. hayatae |
| 10B. |
Sterile flowers strongly curved together. |
13. T. trilobatum |
3.22. KEY TO THE SPECIES OF GENUS ARISAEMA IN VIETNAM
| 1A. |
Leaf blade pedate divided into 3-5 lobes. |
|
| 2A. |
Leaf blade divided into 3 lobes; appendix sessile or not distinguished stipe, slender cylindrical or subulate form, usually with sterile flowers. |
|
| 3A. |
Evergreen rhizomatous plant (Sect.1. Anomala). |
|
| 4A. |
Sterile flowers covered along appendix or only at apex. |
|
| 5A. |
Inflorescence big; appendix long emerged from spathe. |
|
| 6A. |
Spathe with large auricle, lamina without white area at base; appendix green, sterile flower shorter than 2 cm. |
1. A. balansae |
| 6B. |
Spathe without auricle, lamina with white area at base; appendix light brown, sterile flower longer than 2 cm. |
2. A. victoriae |
| 5B. |
Inflorescence small; appendix slightly emerged from spathe tube. |
|
| 7A. |
Sterile flowers only at appendix apex, shorter than 1 cm; spathe with white area at base. |
|
| 8A. |
Rhizome a little woody; stamen without long connection. |
3. A. pingbianense |
| 8B. |
Rhizome not woody; stamens with long connection. |
4. A. rostratum |
| 7B. |
Sterile flowers covered with a long appendix, longer than 1 cm; spathe without white area at lamina base. |
|
| 9A. |
Plant without pseudostem; stamens with short stipe, connection long emerged from anthers |
5. A. omkoiense |
| 9B. |
Plant with pseudostem; stamens with long stipe, connection hidden under anthers |
6. A. petelotii |
| 4B. |
Appendix usually naked or rarely with several sterile flowers at base.(Sect.2. Fimbriata). |
|
| 10A. |
Spathe with large auricle at lamina base |
7. A. garrettii |
| 10B. |
Spathe without auricle. |
|
| 11A. |
Appendix slender cylindrical, blunt at tip. |
8. A. petiolulatum |
| 11B. |
Appendix clavate form, rather stout, round at tip. |
9. A. claviforme |
| 3B. |
Deciduous plant; tuberous. |
|
| 12A. |
Appendix almost naked or with several sterile flowers at base. |
|
| 13A. |
Spathe deep brown, with white area at middle. |
10. A. averyanovii |
| 13B. |
Spathe yellow or white yellow without white area. |
|
| 14A. |
Spathe without auriculate; male spadix with fertile cylindrical, flowers densed |
11. A. pierreanum |
| 14B. |
Auriculate couspicuous; male spadix with fertile portion conical, flowers scattered |
12. A. condaoense |
| 12B. |
Appendix slightly extrosed from spathe tube, with terile flowers at apex or base (Sect.3. Tortuosum). |
|
| 15A. |
Spathe without auricular and with white area at lamina base. |
|
| 16A. |
Appendix with filiforme sterile flowers at apex. |
13. A. ramulosum |
| 16B. |
Appendix without sterile flower at apex. |
14. A. lidaense |
| 15B. |
Spathe with auricular and without white area at lamina base. |
|
| 17A. |
Appendix stout subulate form, soft spine at apex. |
15. A. honbaense |
| 17B. |
Appendix slender subulate, difficult extrude from fathe tube. |
16. A. roxburghii |
| 2B. |
Leaf blade divided in to 5 lobes; appendix with distinct stipe, conical or stout cylindrical, without sterile flowers. |
|
| 18A. |
Evergreen rhizomatous plant; leaf blade divided into 5-7 lobes (Sect.4. Decipientia). |
|
| 19A. |
Spathe purple; appendix elongate conical, truncate at base, acute at apex, straight or bent down as hook, surface smooth. |
17. A. decipiens |
| 19B. |
Spathe yellow-greenish, slightly reddish with black spots; appendix cylindrical, base attenuate, blunt at apex, surface fluted up to apex. |
18. A. rhizomatum
18a. A. rhizomatum var. nudum
|
| 18B. |
Deciduous tuberous plant; leaf blade divided into 3 lobes (Sect.5. Franchetiana) |
19. A. franchetianum |
| 1B. |
Leaf blade radiately divided into 6-15 lobes (Sect.6. Sinarisaema). |
|
| 20A. |
Leaf lobe apices elongated into filiform up to 5 cm long. |
20. A. consanguineum |
| 20B. |
Leaf lobe apices acute or acuminate, c. 1 cm long. |
21. A. parisifolia |
3.23. KEY TO THE SPECIES OF GENUS CRYPTOCORYNE IN VIETNAM
| 1A. |
Spathe longer 10 cm; leaf blade oblong lanceolate to linea form, length at least 5 times width. |
|
| 2A. |
Leaf blade with spots or red stripes, without undulate. |
1. C. crispatula |
| 2B. |
Leaf blade without spots or redstripes, with undulate |
|
| 3B. |
Leaf blade oblong lanceolate, margin nearly straight. |
1a. C. crispatula var. balansae |
| 3A. |
Leaf blade linear form, margin strong waved. |
1b. C. crispatula var. tonkinensis |
| 1B. |
Spathe shorter 10 cm; leaf blade ovate, lanceolate or elliptic, more or less wide, length equal or little longer width |
|
| 4A. |
Spathe lamina twisted. |
2. C. annamica |
| 4B. |
Spathe lamina straight. |
3. C. vietnamensis |
3.24. KEY TO THE SPECIES OF GENUS PISTIA IN VIETNAM
| Floating aquatic plant; leaves spathulate, petioles not conspicuous, rosette in arrangement; spadix bearing only 1 ovary, 2-8 male flowers. |
1. P. stratiotes |









_254903291020251.jpg)






